
RSS-Feed abonnieren
DOI: 10.12945/j.aorta.2014.14-032
Heterogeneity in the Segmental Development of the Aortic Tree
Impact on Management of Genetically Triggered Aortic AneurysmsCorresponding Author
Publikationsverlauf
27. Mai 2014
07. August 2014
Publikationsdatum:
24. September 2018 (online)
- Abstract
- Introduction and Aim of Research
- Literature Search Methodology
- Results
- Discussion
- Conclusions
- References
Abstract
An extensive search of the medical literature examining the development of the thoracic aortic tree reveals that the thoracic aorta does not develop as one unit or in one stage: the oldest part of the thoracic aorta is the descending aorta with the aortic arch being the second oldest, developing under influence from the neural crest cell. Following in chronological order are the proximal ascending aorta and aortic root, which develop from a conotruncal origin. Different areas of the thoracic aorta develop under the influence of different gene sets. These parts develop from different cell lineages: the aortic root (the conotruncus), developing from the mesoderm; the ascending aorta and aortic arch, developing from the neural crest cells; and the descending aorta from the mesoderm. Findings illustrate that the thoracic aorta is not a single entity, in developmental terms. It develops from three or four distinct areas, at different stages of embryonic life, and under different sets of genes and signaling pathways. Genetically triggered thoracic aortic aneurysms are not a monolithic group but rather share a multi-genetic origin. Identification of therapeutic targets should be based on the predilection of certain genes to cause aneurysmal disease in specific aortic segments.
#
“Out of intense complexities, intense simplicities emerge”. —Winston Churchill
Introduction and Aim of Research
Thoracic aortic aneurysms continue to present clinicians with considerable clinical challenges, particularly with risk prediction and its implications on management plan. Previous reports[1] [2] have identified those conditions arising due to inherited causes (i.e., genetically triggered thoracic aortic or GenTAC diseases) as the second most common cause of aortic disease, most frequently clinically recognizable well before the age of 50. Patients affected by such diseases are at a markedly higher risk of mortality (250-fold in some cases) and morbidity, thus making early detection and definitive management imperative.
Current research indicates, however, that these diseases represent a heterogeneous group of conditions[3] [4] sharing only the common feature of “aortic” involvement. Although involvement of the aortic root, aortic annulus, and ascending aorta represents the highest risk to patients, no clear association between any one genetic trigger and a specific area of the thoracic aorta has been definitively established. The fact that GenTAC diseases are usually multigenetic and multifactorial in origin argues against one causative gene or one transcriptional factor being responsible for the disease process in all areas of the thoracic aorta. Even in Marfan syndrome, where there is a global genetic defect of fibrillin maturation not only in the thoracic aorta but also systemically, there is still a marked predilection for the aortic root and proximal ascending aorta, as compared to other aortic segments.
Therefore, we seek to examine (a) whether the thoracic aorta develops as one anatomic unit during one continuous stage, and (b) whether this anatomic development is under the control of one set of genes.
For the purpose of this discussion, the thoracic aorta has been divided into four “fields” ([Fig. 1])—corresponding to their embryologic development—as follows:
-
First aortic field: The descending aorta; from the level of the aortic isthmus (just beyond the left subclavian artery) to the level of the diaphragm.
-
Second aortic field: The aortic arch; from the level of the aortic isthmus to just beyond the origin of the innominate artery.
-
Third aortic field: The ascending aorta; from the sinotubular junction to just before the origin of the innominate artery.
-
Fourth aortic field: The aortic root; from the level of the left ventricular outflow tract to the sino-tubular junction, encompassing the aortic annulus and valve and the earliest portion of the aorta—up to the sinotubular junction.
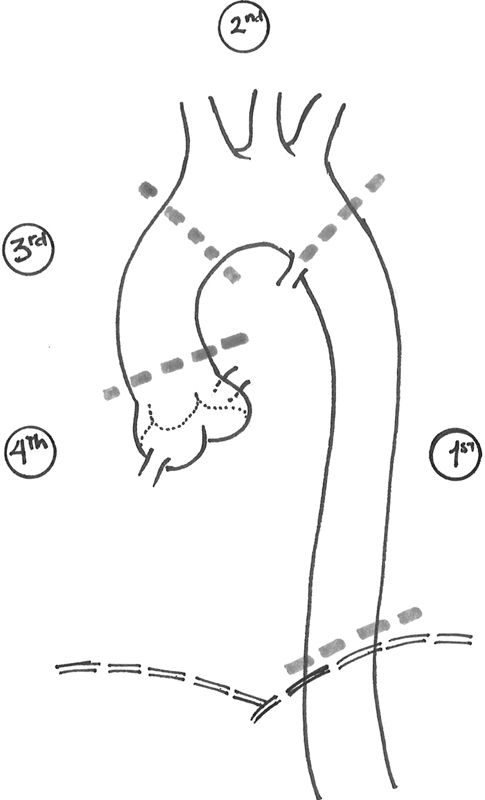
It is important to note that these lines of division do not mark abrupt or sharp transitions but, rather, areas where one area “blends” or “tapers” into the next. This is especially true in the transition between the aortic root and the ascending aorta, around the level of the sinotubular junction.
#
Literature Search Methodology
An extensive search of the medical literature using PubMed, Medline, and Google® Internet search engines, and the National Institutes of Health/National Library of Medicine online databases was performed. Search areas included:
-
Developmental anatomy, folding and looping of the primary heart tube.
-
Development of the different segments of the thoracic aortic tree.
-
Genetics of vascular and aortic development with special focus on segmental effects.
#
Results
The Early Stages and the First Aortic Field (Oldest Segments)
Development of the central arterial vasculature (i.e., the aortic tree) begins quite early, before the initiation of circulation. The first signs of cardiovascular development begin as early as embryonic day (E) 17, with vasculogenesis in two areas on the lateral side of the embryo called “blood islands.” By the early part of the 3rd week (around E19), a pair of vascular elements, called endocardial tubes, is seen. In the early part of the 4th week, as the embryo folds, these two lateral endocardial tubes are brought together in the thoracic region where they fuse to form the primitive heart tube[5] [6] [7] [8] [9] [10] [11] [12].
Around the same time of the endocardial tube development, vascular chords in the mesenchyme of the dorsal body wall form the paired dorsal aortae, both craniad and caudad to the embryonic primitive heart tube. These dorsal aortae also attach to the outflow end of this heart tube ([Fig. 2]).

By E22–E24 (mid-4th week), the primitive heart tube, in turn, undergoes folding along with the generalized axial folding of the embryo. During this process, the endocardial tubes are drawn—along with their attachment to the heart tube—into the ventral aspect of the thorax. At the same time, the part of the paired dorsal aortae attached to the cranial end of the heart tube is pulled ventrally, forming a pair of dorso-ventral “loops,” which are the first aortic arches.
#
The Second Aortic Field (Aortic Arch)
At approximately the same time (beginning E22), mesenchymal cells form five pairs of condensation on either side of the pharyngeal foregut. These five areas correspond to the primitive vertebrate gill bars or branchial arches. Specifically, these five arches in human embryos correspond to branchial arches 1, 2, 3, and 6 in such animals as jawless fish, since the fifth branchial arch either never develops or appears only for a brief period of time before regressing. As mesodermal and endodermal components of these arches are added, their role in human embryos becomes different from that of other animals, thereby giving rise to structures of the lower face, neck, and derivatives of the pharyngeal foregut. Subsequently, these arches in humans are more appropriately termed “pharyngeal arches.”
Development of the pharyngeal arches proceeds in a cranio-caudad order. As a new arch is formed, the aortic sac contributes an artery for that arch. In addition to the first aortic arches described above, the remainder of the vessels associated with the pharyngeal arches (i.e., aortic arches) develop in the ventral aspect from an expansion at the cranial end of the truncus arteriosus called the aortic sac ([Fig. 3]). Aortic arches 2, 3, 4, and 6 develop within their corresponding pharyngeal arches between E26 and E29 (late week 4 to mid-week 5) by a process of vasculogenesis and angiogenesis strongly influenced by migration of neural crest-derived ectomesenchymal cells into these arches.

As the second aortic arch arises by E26, the first aortic arches regress almost completely, without contributing to any mature intrathoracic vasculature. During their regression (around E28), arches 3 and 4 appear. At this stage (Carnegie Stage 13 or approximately E28), the paired dorsal aortae fuse from the level of C7 vertebra to the upper lumbar vertebral level, or at the takeoff of the umbilical artery branch. Finally, the sixth aortic arch forms on E29. Meanwhile, the second aortic arch also regresses without contribution to the intrathoracic vasculature.
By E35 or the end of week 5, segments of the dorsal aorta connecting the third and fourth arches disappear on both sides of the body. This leads to the third arch supplying the head via the cranial extension of the dorsal aorta. Therefore, the third aortic arch gives rise to the right and left common carotid arteries and also the proximal portions of both right and left internal carotid arteries. It is important to keep in mind that the distal internal carotid arteries arise from cranial extensions of the dorsal aorta, while the external carotid arteries develop as outpouching (“sprouts”) from the common carotid arteries.
At a relatively late stage, by week 7, the fourth and sixth arches undergo asymmetrical remodeling to provide the blood supply of the structures in the thoracic inlet and the upper half of the thorax (both upper extremities, both lungs, and the dorsal aorta) At this time, the right-sided dorsal aorta loses its connection with the fused dorsal aortae and the right sixth aortic arch. However, it remains connected to the right fourth arch. It also acquires a branch that later becomes the right subclavian artery, as the area where the right fourth arch connects to the aortic sac becomes modified to become the brachiocephalic (innominate) trunk. Meanwhile, the left fourth arch remains connected to the fused dorsal aortae (the early descending thoracic aortic segment), which, incorporating a small segment of the aortic sac, later becomes the bulk of the aortic arch and the most cranial part of the descending aorta (the area of the aortic isthmus). This means that the remainder of the descending thoracic aorta (from vertebral level T-4 onward) is derived entirely from the older fused dorsal aortae without any contribution from the aortic arches or other segments ([Fig. 4]).

Development of the sixth aortic arches is dramatically asymmetric. By late week 7, and as the distal right sixth arch loses its connection with the right dorsal aorta and disappears, the left sixth arch persists to later become the ductus arteriosus, connecting to the distal end of the definitive aortic arch[12] [13] [14] [15] [16] [17] [18] [19] [20].
#
The Third Aortic Field (Ascending Aorta)
After the primitive heart tube is formed, several sulci appear around early week 4 to divide it into distinct chambers/structures. The inferior, caudad or inflow (systemic venous) end describes the sinus venosus. Next as we move cranially, the next two chambers (atrium and ventricle) are separated by the atrioventricular sulcus. In turn, the ventricle is separated from the bulbus cordis by the bulboventricular sulcus. The most cranial part of the bulbus is called the conotruncus, whose distal part connects to a dilated expansion called aortic sac, which is continuous with the first aortic arches, and to which the third, fourth, and sixth arches also connect as previously discussed.
After the heart tube undergoes folding by week 5, septation of the ventricle commences. Starting around the same time, right and left trunco-conal swellings grow out from the lateral walls of the common ventricular outflow tract. As they meet, these swellings would grow superiorly and inferiorly inside the outflow tract, dividing it in a spiral fashion ([Fig. 5]). By week 9, these would have grown into the upper ridge of the muscular interventricular septum and thus completed the separation between the aortic and pulmonary outflow tracts[21] [22] [23] [24] [25] [26].

#
The Fourth Aortic Field (Aortic Root)
At the time of emergence of the truncal swellings, a second pair of tubercles arises at the anterior and posterior walls of the truncus at the same level. Thus, the anteriorly located tubercle is situated in the pulmonary channel after the septation is completed, while the posteriorly located one will be in the aortic channel. Toward the end of week 9, and after septation has been completed, each outflow tract will contain a triangle of tubercles: two from the older lateral wall tubercles and the third from either the anterior or posterior wall. In each channel, this triangle will give rise to the three cusps of the aortic and pulmonary semilunar valves. As such, the aortic valve and very proximal part of the ascending aorta (i.e., the aortic root) develop as a result of the septation of the distal aspect of the conotruncus, which is an essential component of the primitive heart tube[27] [28] [29] [30] [31] [32].
#
Genetic and Molecular Mechanisms
(a) Development of the dorsal aorta is influenced by multiple genes and signaling pathways. Vascular endothelial growth factor A (VEGF-A) is activated by sonic hedge hog (Shh) molecule and, in turn, activates the Notch signaling pathway through its ligand δ-like α, VGEFR, and phospholipase-C1 enzyme (PLCγ-1). Several other genes are involved, such as Rbm 24; SMAD5, which are downstream signal regulators of the TGF-β receptors; Sox-13 gene (Sry-related HMG box); Sox-18 transcription factor; Hox genes; hypoxia-inducible factor 1α (HIF-1α); zinc finger protein genes; and cyclo-oxygenase-1 (Cox-1)-derived prostaglandins under the Flil promoter.
In addition, other vertebrate-specific chemokines such as the C-X-C motif cxcra and its ligand cxcl12b are involved in development of the lateral dorsal aorta and are required for the ventral migration of the endoderm-derived parts. Thus, the anterior part of the each of the paired lateral dorsal aortae migrates ventrally under the influence of Kdrl, cxcra, and Flil. Since the posterior part of the dorsal aorta lacks Kdrl, it maintains its anatomic position next to the spine. This process of ventral migration of the first (anterior) parts (forerunners of the first aortic arches and the aortic sac) is also influenced by the increased regulators of the G protein signaling via S1P1, S1P2, S1P3, and Fibulin-1 expression at the ventral aspects of the dorsal aorta.
Other genes that are primarily involved in the formation of the primary heart tube include MesP1, β-galactosidase gene (lac-Z), canonical and noncanonical Wnt signaling of the Frizzled (FZD) gene and β-catenin, Nk2.5, GATA4, and Tbx2 and Tbx3 [33] [34] [35] [36] [37] [38] [39] [40] [41].
(b) Genetic and molecular control of the pharyngeal arches is rather complex and involves a number of concurrent processes. First is the migration of ectoderm-derived neural crest cells ventrally to the area of the arch arteries. This is regulated by Tbx1 which regulates Gbx2 and bone-morphogenic protein Bmp4 expression in the pharyngeal ectoderm. Lbx1 homeobox gene and Pax3 are also implicated in the migration of a specific subpopulation of neural crest cells. The specific sequential patterning of the arches is under the influence of multiple factors: Wnt1 gene is involved in the dorso-ventral patterning and is also expressed at the time of neural crest cell migration. Pitx2c, a left-right differentiation gene, is expressed in the left aortic sac and at the junction of the aortic sac and branchial arches. It plays a role in the asymmetric branchial arch development. This effect seems to be also influenced by the difference in blood flow via platelet-derived growth factor (PDGF), and vascular endothelial growth factor (VEGF) patterning of the aortic arches, particularly the fourth arch, involves both Fgf8 and Tbx1 genes; however, the effect of Fgf8 is limited to the aortic arch and not the outflow tract (conotruncus). There also seems to be a synergistic effect between Tbx1 and Chd7 genes. Frizzled (FZD2) gene appears in the pharyngeal mesenchyme around E10, then its expression decreases until E18 when it is only seen in the aorta and pulmonary trunk. Hox gene homologues are responsible for providing each segment of the pharyngeal arches with its positional information, thus acquiring its own identity. Prx1(MHox) and Prx2(S8) are both nonclustered homeobox genes involved in the architecture of the great vessels and the ductus arteriosus. Hoxa genes are involved especially in the third arch. There is a synergy between Hoxa1 and Hoxb1 in arch patterning and generation of the cranial neural crest, while Hoxa2 and Hoxa3 are involved in the stability of patterning. The eHAND gene and its dHAND transcription factor are involved in regulating the branchial arch mesenchyme.
Other factors include endothelin (Et-1 or Edn-1) and its endothelin-converting enzyme (Ece-1) signaling, which involves G protein-coupled receptors, independent of SM22 and lac-Z. Tumor growth factor β (TGFβ-1) via Emad2 and fibronectin influences the fourth arch. Smad2 expression is not observed in other arch segments, the ascending or descending aorta. TFAP2B gene and the encoding transcription factor activating enhancer-binding protein 2β (AP2β) have been linked to persistent ductus arteriosus families and the Char syndrome. JAG1, GJA 1 (connexin 43), and Ednra genes have been implicated in Algille syndrome and coarctation of the aorta, hypoplastic left heart syndrome, familial aortic aneurysms, and patent ductus arteriosus, respectively.
(c) Outflow Tract and the Conotruncus: Cardiac neural crest cells migrate from the area of the 4th arch under the influence of their drivers Wnt1-cre and Pax3 cre , along with histone deacetylase 3 (Hdac3). Tbx1 is required for the growth, alignment and septation of the outflow tract. It regulates Fgf8 which is involved in differentiation of the arterial pole. Septation is also dependent on the expression of Madh6 genes, encoding for Smad6 protein, a signaling protein involved with the TGF-β superfamily, as well as the Frizzled 2 (FZD2) gene and the HIRA gene on chromosome 22. Other factors include Bmp2, Nkx2.5, Pitx2, Fibroblast growth factor (Fgf). Fgf15 is expressed transiently in the aortic arches (D9.5 to D10), but its absence causes failure of the aorta to wedge between the tricuspid and mitral valves. It also affects the proximal outflow tract at level of its connection with the 6th arch. Prominent levels of tropoelastin (TE) are seen in the aorta and pulmonary trunk. Thyroid hormone-receptor associated protein-2 (THARP2) gene mutations have been implicated in the genesis of transposition of the great vessels. Defects in the human homologues of dishevelled gene and β-catenin are associated with conotruncal defects[42] [43] [44] [45] [46] [47] [48] [49] [50] [51] [52] [53] [54] [55] [56] [57].
(d) The aortic root and proximal ascending aorta. Although this area of the aortic tree carries a high clinical significance, there is a comparative paucity with regard to identification of the genetic and molecular factors influencing its development and differentiation. So far, the following genes have been implicated in proximal aortic pathology: Notch1 and Sirtuin1 (SirT1)—a longevity gene limiting Notch1 signaling—have both been associated with bicuspid aortic valve. Abnormalities of chromosomes 45, 13, 18, X, and 21 have all been linked to various conditions including Turner syndrome, hypoplastic left heart syndrome, and bicuspid aortic valve. FLNA (flamin) gene and the Ras-mitogen-associated protein kinase signaling pathway (SHP-2, ki-Ras, RAF-1, and SOS-1) have also been implicated in the same condition. Conversely, defects or deletions of the elastin gene on chromosome 7, also in association with Wnt-Frizzled homologue (FZD9), results in supravalvular aortic stenosis[58] [59] [60] [61] [62] [63] [64] [65] [66] [67] [68] [69] [70] [71] [72] [73] [74] [75] [76] [77] [78] [79] [80] [81] [82] [83].
#
#
Discussion
From a developmental standpoint, the thoracic aorta is not a uniform, homogenous structure. It does not describe one contiguous anatomic entity, nor do all its segments develop at the same time. In chronologic terms, the thoracic aortic tree can be divided into the above-mentioned four “fields” (see above), each developing at a different stage and from a different cell population ancestry. As elaborated above, the oldest part of the thoracic aorta is the descending aorta distal to the isthmus (T-4 level), with the aortic arch following in chronological order. The distal ascending aorta develops next, followed by the aortic root and proximal ascending aorta. Each of these “aortic fields” develops under the influence of a distinct set of genes and signaling molecules and pathways.
In concurrence with the differences between the thoracic and abdominal aortic regions previously described[84], our review further elaborates on the different embryologic origin, genetic influence, and developmental trajectory of areas with the thoracic aorta itself. Despite the recent emphasis[85] [86] on genetic and signaling pathways involved in the vascular smooth muscle development and maturation, which are seen as exerting a global influence over the entire vascular tree, a single gene, receptor, or signaling pathway does not seem to have a generalized effect on the development of specific segments of the aortic tree. As an example, TGF-β effect on the smooth muscle cell gene expression differs depending on the location of the smooth muscle cell in the aorta. TGF-β isoforms have different effects on the smooth muscle transcriptional response in a lineage-dependent manner, with the highest response in lineage of ectodermal origin (the outflow tract and proximal ascending aorta) versus the lowest response in mesodermal lineage (dorsal aorta). The same applies to other genes including smooth muscle M α-actin, SM-myosin heavy chain (MYH-11), SM22α and other SMC proteins (lysyl oxidase, fibulin 4, fibulin 5), and tropoelastin. This further highlights the heterogenicity of the genetic and signaling factors behind the development of each segment (field) of the thoracic aorta. These findings also have an increased significance considering the role of such factors in the regulation of the inflammatory response, atherogenesis, and the extracellular matrix, all of which contribute to the structural integrity of the aortic wall.
#
Conclusions
In developmental terms, the thoracic aorta is a complex, heterogeneous structure. No single gene, transcription factor, or molecule regulates the entire process of aortic development. Each segment of the aorta develops and differentiates under a distinct set of genetic and transcriptional factors. Therefore, it is inappropriate to speak of “thoracic aortic aneurysm” as a monolithic entity, or of any single cause as responsible for all types and locations of aneurysmal disease. In addition, this heterogeneity may suggest that the areas where these different thoracic aortic segments join are under complex and perhaps conflicting genetic, developmental, and regulatory influence. As such, these areas of juncture (the “weld joints”) may be at a higher risk for anomalies in development, differentiation, connection, or functional regulation, thereby rendering them more susceptible to disease processes.
Registry-based research has emerged recently as a platform for robust investigation utilizing fairly large amounts of real-world population data organized in condition-specific databases. Another advantage is the coordinated efforts between different teams of researchers across several disciplines. The GenTAC National Registry[87] has been established by a collaborative effort between the US Department of Health and Human Services and the National Institutes of Health to help healthcare providers better understand the links between genes and thoracic aortic and cardiovascular disease. This registry is a repository for data concerning the genetics, anatomy, pathophysiology, and natural history of thoracic aortic conditions. As such, it is uniquely positioned to organize and undertake the methodical classification of each individual condition, in terms of its genetic etiology and the transcriptional and signaling pathways and specific anatomic locale along the thoracic aorta affected. The identification of which areas of the thoracic aortic tree are most commonly affected in which genetic and/or translational or signaling defect would thus provide a sound scientific basis for formulating and implementing disease-specific testing, screening, and therapeutic modalities. This is fundamental to the development of evidence-based, specific practice guidelines, which will translate into improved effectiveness, safety, and efficiency of the management of thoracic aortic disease in this challenging group of patients.
#
#
Conflict of Interest
The authors have no conflict of interest relevant to this publication.
-
References
- 1 Caglayan AO, Dundar M. Inherited diseases and syndromes leading to aortic aneurysms and dissections. Eur J Cardiothorac Surg 2009; 35: 931-940 . 10.1016/j.ejcts.2009.01.006
- 2 Jondeau G. Phenotypic heterogeneity and determinants in aortic aneurysm syndromes. . Proceedings, the Aortic Disease Summit. Baltimore, Maryland. September 22-23, 2009
- 3 Elefteriades JA, Farkas EA. Thoracic aortic aneurysm clinically pertinent controversies and uncertainties. J Am Coll Cardiol 2010; 55: 841-857 . 10.1016/j.jacc.2009.08.084
- 4 Sherif HMF. In search of a new therapeutic target for the treatment of genetically triggered thoracic aortic aneurysms and cardiovascular conditions: insights from human and animal lathyrism. Interact CardioVasc Thorac Surg 2010; 11: 271-276 . 10.1510/icvts.2010.239681
- 5 Winstein B. What guides early embryonic blood vessel formation?. Dev Dyn 1999; 215: 2-11
- 6 Larsen W.. Development of the heart. In: Human Embryology. New York: Churchill Livingstone; 1997. , p. 135-161
- 7 Abu-Issa R, Kirby ML. Heart field: from mesoderm to heart tube. Annu Rev Cell Dev Biol 2007; 23: 45-68 . 10.1146/annurev.cellbio.23.090506.123331
- 8 Siekmann AF, Standley C, Fogarty KE, Wolfe SA, Lawson ND. Chemokine signaling guides regional patterning of the first embryonic artery. Genes Dev 2009; 23: 2272-2277 . 10.1101/gad.1813509
- 9 Cleaver, Krieg P. VEGF mediates angioblast migration during development of the dorsal aorta in Xenopus. Development 1998; 125: 3905-3914
- 10 McLean SE, Mecham BH, Kelleher CM, Mariani TJ, Mecham RP. Extracellular matrix gene expression in the developing mouse aorta. Adv Dev Biol 2005; 15: 81-128 . 10.1016/S1574-3349(05)15003-0
- 11 Hungerford JE, Owens GK, Argraves WS, Little CD. Development of the aortic vessel wall as defined by vascular smooth muscle and extracellular matrix markers. Dev Biol 1996; 178: 375-392 . 10.1006/dbio.1996.0225
- 12 Eriksson J, Löfberg J. Development of the hypochord and dorsal aorta in the zebrafish embryo (Danio rerio). J Morphol 2000; 244: 167-176 . 10.1002/(SICI)1097-4687(200006)244:3<167::AID-JMOR2>3.0.CO;2-J
- 13 Hunt P, Clarke JD, Buxton P, Ferretti P, Thorogood P. Stability and plasticity of neural crest patterning and branchial arch Hox code after extensive cephalic crest rotation. Dev Biol 1998; 198: 82-104 . 10.1006/dbio.1998.8886
- 14 Bockman DE, Redmond ME, Kirby ML. Alteration of early vascular development after ablation of cranial neural crest. Anat Rec 1989; 225: 209-217 . 10.1002/ar.1092250306
- 15 Bockman DE, Redmond ME, Waldo K, Davis H, Kirby ML. Effect of neural crest ablation on development of the heart and arch arteries in the chick. Am J Anat 1987; 180: 332-341 . 10.1002/aja.1001800403
- 16 Bergwerff M, DeRuiter MC, Hall S, Poelmann RE, Gittenberger-de Groot AC. Unique vascular morphology of the fourth aortic arches: possible implications for pathogenesis of type-B aortic arch interruption and anomalous right subclavian artery. Cardiovasc Res 1999; 44: 185-196 . 10.1016/S0008-6363(99)00186-8
- 17 Kirby ML. Cellular and molecular contributions of the cardiac neural crest to cardiovascular development. Trends Cardiovasc Med 1993; 3: 18-23 . 10.1016/1050-1738(93)90023-Y
- 18 Porras D, Brown CB. Temporal-spatial ablation of neural crest in the mouse results in cardiovascular defects. Dev Dyn 2008; 237: 153-162 . 10.1002/dvdy.21382
- 19 Kirby ML, Hunt P, Wallis K, Thorogood P. Abnormal patterning of the aortic arch arteries does not evoke cardiac malformations. Dev Dyn 1997; 208: 34-47 . 10.1002/(SICI)1097-0177(199701)208:1<34::AID-AJA4>3.0.CO;2-2
- 20 Yashiro K, Shiratori H, Hamada H. Haemodynamics determined by a genetic programme govern asymmetric development of the aortic arch. Nature 2007; 450: 285-288 . 10.1038/nature06254
- 21 Larsen W.. Development of the heart. In: Human Embryology. New York: Churchill Livingstone; 1997. , p. 169-187
- 22 Bruneau B. The developmental genetics of congenital heart disease. Nature 2008; 451: 943-948 . 10.1038/nature06801
- 23 Nishibatake M, Kirby ML, Van Mierop LH. Pathogenesis of persistent truncus arteriosus and dextroposed aorta in the chick embryo after neural crest ablation. Circulation 1987; 75: 255-264 . 10.1161/01.CIR.75.1.255
- 24 Hutson MR, Kirby ML. Model systems for the study of heart development and disease. Cardiac neural crest and conotruncal malformations. Semin Cell Dev Biol 2007; 18: 101-110 . 10.1016/j.semcdb.2006.12.004
- 25 Restivo A, Piacintini G, Placidi S, Saffirio C, Marino B. Cardiac outflow tract: a review of some embryogenetic aspects of the conotruncal region of the heart. Anat Rec A Discov Mol Cell Evol Biol 2006; 288: 936-943 . 10.1002/ar.a.20367
- 26 Beall AC, Rosenquist TH. Smooth muscle cells of neural crest origin form the aorticopulmonary septum in the avian embryo. Anat Rec 1990; 226: 360-366 . 10.1002/ar.1092260313
- 27 Kappetein A, Gittenberger-de Groot A, Zwinderman A, Rohmer J, Poelmann R, Huysmans H. The neural crest as a possible pathogenetic factor in coarctation of the aorta and bicuspid aortic valve. J Thorac Cardiovasc Surg 1991; 102: 830-836
- 28 Anderson RH, Webb S, Brown NA, Lamers W, Moorman A. Development of the heart: (3) formation of the ventricular outflow tracts, arterial valves, and intrapericardial arterial trunks. Heart 2003; 89: 1110-1118 . 10.1136/heart.89.9.1110
- 29 Qayyum SR, Webb S, Anderson RH, Verbeek FJ, Brown NA, Richardson MK. Septation and valvar formation in the outflow tract of the embryonic chick heart. Anat Rec 2001; 264: 273-283 . 10.1002/ar.1162
- 30 Webb S, Qayyum SR, Anderson RH, Lamers WH, Richardson MK. Septation and separation within the outflow tract of the developing heart. J Anat 2003; 202: 327-342 . 10.1046/j.1469-7580.2003.00168.x
- 31 Maron B, Hutchins G. The development of the semilunar valves in the human heart. Am J Pathol 1974; 74: 333-344
- 32 Fernández B, Durán AC, Fernández-Gallego T, Fernández MC, Such M, Arqué JM. , et al. Bicuspid aortic valves with different spatial orientations of the leaflets are distinct etiologic entities. J Am Coll Cardiol 2009; 54: 2312-2318 . 10.1016/j.jacc.2009.07.044
- 33 Cho H, Harrison K, Schwartz O, Kehrl JH. The aorta and heart differentially express RGS (regulators of G-protein signalling) proteins that selectively regulate sphingosine 1-phosphate, angiotensin II and endothelin-1 signalling. Biochem J 2003; 371: 973-980 . 10.1042/BJ20021769
- 34 Downes M, Koopman P. SOX18 and the transcriptional regulation of blood vessel development. Trends Cardiovasc Med 2001; 11: 318-324 . 10.1016/S1050-1738(01)00131-1
- 35 Yang X, Castilla LH, Xu X, Li C, Gotay J, Weinstein M. , et al. Angiogenesis defects and mesenchymal apoptosis in mice lacking SMAD5. Development 1999; 126: 1571-1580
- 36 Zhong TP, Rosenberg M, Mohideen MA, Weinstein B, Fishman MC. gridlock, an HLH gene required for assembly of the aorta in zebrafish. Science 2000; 287: 1820-1824 . 10.1126/science.287.5459.1820
- 37 Jaffe M, Sesti C, Washington IM, Du L, Dronadula N, Chin MT. , et al. Transforming growth factor β signaling in myogenic cells regulates vascular morphogenesis, differentiation and matrix synthesis. Arterioscler Thromb Vasc Biol 2012; 23: e1-e11 . 10.1161/ATVBAHA.111.238410
- 38 Topouzis S, Majesky M. Smooth muscle lineage diversity in the chick embryo: two types of aortic smooth muscle cell differ in growth and receptor-mediated transcriptional responses to transforming growth factor-β. Dev Biol 1996; 178: 430-445 . 10.1006/dbio.1996.0229
- 39 Patterson LJ, Gering M, Patient R. Scl is required for dorsal aorta as well as blood formation in zebrafish embryos. Blood 2005; 105: 3502-3511 . 10.1182/blood-2004-09-3547
- 40 Cha YI, Kim SH, Solnica-Krezel L, DuBois RN. Cyclooxygenase-1 signaling is required for vascular tube formation during development. Dev Biol 2005; 282: 274-283 . 10.1016/j.ydbio.2005.03.014
- 41 Ben Abdelkhalek H, Beckers A, Schuster-Gossler K, Pavlova MN, Burkhardt H, Lickert H. , et al. The mouse homeobox gene Not is required for caudal notochord development and affected by the truncate mutation. Genes Dev 2004; 18: 1725-1736 . 10.1101/gad.303504
- 42 Kuo CT, Morrisey EE, Anandappa R, Sigrist K, Lu MM, Parmacek MS. , et al. GATA4 transcription factor is required for ventral morphogenesis and heart tube formation. Genes Dev 1997; 11: 1048-1060 . 10.1101/gad.11.8.1048
- 43 Holzenberger M, Ayer-Le Lièvre, Robert L. Tropoelastin gene expression in the developing vascular system of the chicken: an in situ hybridization study. Anat Embryol 1993; 188: 481-492
- 44 Abdulla RI, Slott EF, Kirby ML. Proteins associated with cardiac neural crest in the pharyngeal region of early chick embryos. Pediatr Res 1993; 33: 43-47 . 10.1203/00006450-199301000-00009
- 45 Biben C, Hatzistavrou T, Harvey R. Expression of NK-2 class homeobox gene Nkx2-6 in foregut endoderm and heart. Mech Dev 1998; 73: 125-127 . 10.1016/S0925-4773(98)00037-9
- 46 Calmont A, Ivins S, Van Bueren KL, Papangeli I, Kyriakopoulou V, Andrews WD. , et al. Tbx1 controls cardiac neural crest cell migration during arch artery development by regulating Gbx2 expression in the pharyngeal ectoderm. Development 2009; 136: 3173-3183 . 10.1242/dev.028902
- 47 Gittenberger-de Groot AC, Azhar M, Molin DG. Transforming growth factor beta-SMAD2 signaling and aortic arch development. Trends Cardiovasc Med 2006; 16: 1-6 . 10.1016/j.tcm.2005.09.006
- 48 Waldo KL, Lo CW, Kirby ML. Connexin 43 expression reflects neural crest patterns during cardiovascular development. Dev Biol 1999; 208: 307-323 . 10.1006/dbio.1999.9219
- 49 Yanagisawa H, Hammer RE, Richardson JA, Williams SC, Clouthier DE, Yanagisawa M. Role of endothelin-1/endothelin-A-receptor-mediated signaling pathway in the aortic arch patterning in mice. J Clin Invest 1998; 102: 22-33 . 10.1172/JCI2698
- 50 Morishima M, Yanagisawa H, Yanagisawa M, Baldini A. Ece1 and Tbx1 define distinct pathways to aortic arch morphogenesis. Dev Dyn 2003; 228: 95-104 . 10.1002/dvdy.10358
- 51 Gavalas A, Trainor P, Ariza-McNaughton L, Krumlauf R. Synergy between Hoxa1 and Hoxb1: the relationship between arch patterning and the generation of cranial neural crest. Development 2001; 128: 3017-3027
- 52 Kameda Y. Hoxa3 and signaling molecules involved in aortic arch patterning and remodeling. Cell Tissue Res 2009; 336: 165-178 . 10.1007/s00441-009-0760-7
- 53 Liao J, Aggarwal VS, Nowotschin S, Bondarev A, Lipner S, Morrow BE. Identification of downstream genetic pathways of Tbx1 in the second heart field. Dev Biol 2008; 316: 524-537 . 10.1016/j.ydbio.2008.01.037
- 54 Molin DG, Poelmann RE, DeRuiter MC, Azhar M, Doetschman T, Gittenberger-de Groot AC. Transforming growth factor beta-SMAD2 signaling regulates aortic arch innervation and development. Circ Res 2004; 95: 1109-1117 . 10.1161/01.RES.0000150047.16909.ab
- 55 Ohnemus S, Kanzler B, Jerome-Majewska L, Papaioannou VE, Boehm T, Mallo M. Aortic arch and pharyngeal phenotype in the absence of BMP-dependent neural crest in the mouse. Mech Dev 2002; 119: 127-135 . 10.1016/S0925-4773(02)00345-3
- 56 Schäfer K, Neuhaus P, Kruse J, Braun T. The homeobox gene Lbx1 specifies a subpopulation of cardiac neural crest necessary for normal heart development. Circ Res 2002; 92: 73-80 . 10.1161/01.RES.0000050587.76563.A5
- 57 Thomas T, Kurihara H, Yamagishi H, Kurihara Y, Yazaki Y, Olson EN. , et al. A signaling cascade involving endothelin-1, dHAND and msx1 regulates development of neural-crest-derived branchial arch mesenchyme. Development 1998; 125: 3005-3014
- 58 Hutson MR, Zhang P, Stadt HA, Sato AK, Li YX, Burch J. , et al. Cardiac arterial pole alignment is sensitive to FGF8 signaling in the pharynx. Dev Biol 2006; 295: 486-497 . 10.1016/j.ydbio.2006.02.052
- 59 van Gijn M, Blankesteijn W, Smits J, Hierck B, Gittenberger-de Groot A. Frizzled 2 is transiently expressed in neural crest-containing areas during development of the heart and great arteries in the mouse. Anat Embryol 2001; 203: 185-192
- 60 Vitelli F, Zhang Z, Huynh T, Sobotka A, Mupo A, Baldini A. Fgf8 expression in the Tbx1 domain causes skeletal abnormalities and modifies the aortic arch but not the outflow tract phenotype of Tbx1 mutants. Dev Biol 2006; 295: 559-570 . 10.1016/j.ydbio.2006.03.044
- 61 Vitelli F, Taddei I, Morishima M, Meyers EN, Lindsay EA, Baldini A. A genetic link between Tbx1 and fibroblast growth factor signaling. Development 2002; 129: 4605-4611
- 62 Randall V, McCue K, Roberts C, Kyriakopoulou V, Beddow S, Barrett AN. , et al. Great vessel development requires biallelic expression of Chd7 and Tbx1 in pharyngeal ectoderm in mice. J Clin Invest 2009; 119: 3301-3310 . 10.1172/JCI37561
- 63 Scambler PJ. 22q11 deletion syndrome: a role for TBX1 in pharyngeal and cardiovascular development. Pediatr Cardiol 2010; 31: 378-390 . 10.1007/s00246-009-9613-0
- 64 Bergwerff M, Gittenberger-de Groot AC, Wisse LJ, DeRuiter MC, Wessels A, Martin JF. , et al. Loss of function of the Prx1 and Prx2 homeobox genes alters architecture of the great elastic arteries and ductus arteriosus. Virchows Arch 2000; 436: 12-19 . 10.1007/PL00008193
- 65 Farrell MJ, Stadt H, Wallis KT, Scambler P, Hixon RL, Wolfe R. , et al. HIRA, a DiGeorge syndrome candidate gene, is required for cardiac outflow tract septation. Circ Res 1999; 84: 127-135 . 10.1161/01.RES.84.2.127
- 66 Heathcote K, Braybrook C, Abushaban L, Guy M, Khetyar ME, Patton MA. , et al. Common arterial trunk associated with a homeodomain mutation of NKX2.6. Hum Mol Genet 2005; 14: 585-593 . 10.1093/hmg/ddi055
- 67 Mesbah K, Harrelson Z, Théveniau-Ruissy M, Papaioannou VE, Kelly RG. Tbx3 is required for outflow tract development. Circ Res 2008; 103: 743-750 . 10.1161/CIRCRESAHA.108.172858
- 68 Nie X, Brown CB, Wang Q, Jiao K. Inactivation of Bmp4 from the Tbx1 expression domain causes abnormal pharyngeal artery and cardiac outflow tract remodeling. Cells Tissue Organs 2011; 193: 393-403 . 10.1159/000321170
- 69 Washington Smoak I, Byrd NA, Abu-Issa R, Goddeeris MM, Anderson R, Morris J. , et al. Sonic hedgehog is required for cardiac outflow tract and neural crest cell development. Dev Biol 2005; 283: 357-372 . 10.1016/j.ydbio.2005.04.029
- 70 Kim RY, Robertson EJ, Solloway MJ. Bmp6 and Bmp7 are required for cushion formation and septation in the developing mouse heart. Dev Biol 2001; 235: 449-466 . 10.1006/dbio.2001.0284
- 71 Xu H, Morishima M, Wylie JN, Schwartz RJ, Bruneau BG, Lindsay EA. , et al. Tbx1 has a dual role in the morphogenesis of the cardiac outflow tract. Development 2004; 131: 3217-3227 . 10.1242/dev.01174
- 72 Vincentz JW, McWhirter JR, Murre C, Baldini A, Furuta Y. Fgf15 is required for proper morphogenesis of the mouse cardiac outflow tract. Genesis 2005; 41: 192-201 . 10.1002/gene.20114
- 73 Chen YH, Ishii M, Sun J, Sucov HM, Maxson Jr RE. Msx1 and Msx2 regulate survival of secondary heart field precursors and post-migratory proliferation of cardiac neural crest in the outflow tract. Dev Biol 2007; 308: 421-437 . 10.1016/j.ydbio.2007.05.037
- 74 Liao J, Aggarwal VS, Nowotschin S, Bondarev A, Lipner S, Morrow BE. Identification of downstream genetic pathways of Tbx1 in the second heart field. Dev Biol 2008; 316: 524-537 . 10.1016/j.ydbio.2008.01.037
- 75 Parisot P, Mesbah K, Théveniau-Ruissy M, Kelly RG. Tbx1, subpulmonary myocardium and conotruncal congenital heart defects. Birth Defects Res A Clin Mol Teratol 2011; 91: 477-484 . 10.1002/bdra.20803
- 76 Raid R, Krinka D, Bakhoff L, Abdelwahid E, Jokinen E, Kärner M. , et al. Lack of Gata3 results in conotruncal heart anomalies in mouse. Mech Dev 2009; 126: 80-89 . 10.1016/j.mod.2008.10.001
- 77 Jiang X, Rowitch DH, Soriano P, McMahon AP, Sucov HM. Fate of the mammalian cardiac neural crest. Development 2000; 127: 1607-1616
- 78 Hove JR, Köster RW, Forouhar AS, Acevedo-Bolton G, Fraser SE, Gharib M. Intracardiac fluid forces are an essential epigenetic factor for embryonic cardiogenesis. Nature 2003; 421: 172-177 . 10.1038/nature01282
- 79 Ewart AK, Morris CA, Ensing GJ, Loker J, Moore C, Leppert M. , et al. A human vascular disorder, supravalvular aortic stenosis, maps to chromosome 7. Proc Natl Acad Sci U S A 1993; 90: 3226-3230 . 10.1073/pnas.90.8.3226
- 80 Galvin KM, Donovan MJ, Lynch CA, Meyer RI, Paul RJ, Lorenz JN. , et al. A role for Smad6 in development and homeostasis of the cardiovascular system [Letter]. Nat Genet 2000; 24: 171-174 . 10.1038/72835
- 81 Martin LJ, Ramachandran V, Cripe LH, Hinton RB, Andelfinger G, Tabangin M. , et al. Evidence in favor of linkage to human chromosomal regions 18q, 5q and 13q for bicuspid aortic valve and associated cardiovascular malformations. Hum Genet 2007; 121: 275-284 . 10.1007/s00439-006-0316-9
- 82 Sans-Coma V, Cardo M, Durán AC, Franco D, Fernández B, Arqué JM. Evidence for quantitative genetic influence on the formation of aortic valves with 2 leaflets in the Syrian hamster. Cardiol Young 1993; 3: 132-140
- 83 Olson TM, Michels VV, Urban Z, Csiszar K, Christiano AM, Driscoll DJ. , et al. A 30kb deletion within the elastin gene results in familial supravalvular aortic stenosis. Hum Mol Genet 1995; 9: 1677-1679
- 84 Ruddy JM, Jones JA, Spinale FG, Ikonomidis JS. Regional heterogeneity within the aorta: relevance to aneurysm disease. J Thorac Cardiovasc Surg 2008; 136: 1123-1130 . 10.1016/j.jtcvs.2008.06.027
- 85 Milewicz DM, Guo DC, Tran-Fadulu V, Lafont AL, Papke CL, Inamoto S. , et al. Genetic basis of thoracic aortic aneurysms and dissections: focus on smooth muscle cell contractile dysfunction. Annu Rev Genom Hum Genet 2008; 9: 283-302 . 10.1146/annurev.genom.8.080706.092303
- 86 Wang L, Guo DC, Cao J, Gong L, Kamm KE, Regalado E. , et al. Mutations in myosin light chain kinase cause familial aortic dissections. Am J Hum Genet 2010; 87: 701-707 . 10.1016/j.ajhg.2010.10.006
- 87 The National Registry of Genetically Triggered Thoracic Aortic Aneurysms and Cardiovascular Conditions (GenTAC). Available online at: https://gentac.rti.org/Home.aspx
Corresponding Author
-
References
- 1 Caglayan AO, Dundar M. Inherited diseases and syndromes leading to aortic aneurysms and dissections. Eur J Cardiothorac Surg 2009; 35: 931-940 . 10.1016/j.ejcts.2009.01.006
- 2 Jondeau G. Phenotypic heterogeneity and determinants in aortic aneurysm syndromes. . Proceedings, the Aortic Disease Summit. Baltimore, Maryland. September 22-23, 2009
- 3 Elefteriades JA, Farkas EA. Thoracic aortic aneurysm clinically pertinent controversies and uncertainties. J Am Coll Cardiol 2010; 55: 841-857 . 10.1016/j.jacc.2009.08.084
- 4 Sherif HMF. In search of a new therapeutic target for the treatment of genetically triggered thoracic aortic aneurysms and cardiovascular conditions: insights from human and animal lathyrism. Interact CardioVasc Thorac Surg 2010; 11: 271-276 . 10.1510/icvts.2010.239681
- 5 Winstein B. What guides early embryonic blood vessel formation?. Dev Dyn 1999; 215: 2-11
- 6 Larsen W.. Development of the heart. In: Human Embryology. New York: Churchill Livingstone; 1997. , p. 135-161
- 7 Abu-Issa R, Kirby ML. Heart field: from mesoderm to heart tube. Annu Rev Cell Dev Biol 2007; 23: 45-68 . 10.1146/annurev.cellbio.23.090506.123331
- 8 Siekmann AF, Standley C, Fogarty KE, Wolfe SA, Lawson ND. Chemokine signaling guides regional patterning of the first embryonic artery. Genes Dev 2009; 23: 2272-2277 . 10.1101/gad.1813509
- 9 Cleaver, Krieg P. VEGF mediates angioblast migration during development of the dorsal aorta in Xenopus. Development 1998; 125: 3905-3914
- 10 McLean SE, Mecham BH, Kelleher CM, Mariani TJ, Mecham RP. Extracellular matrix gene expression in the developing mouse aorta. Adv Dev Biol 2005; 15: 81-128 . 10.1016/S1574-3349(05)15003-0
- 11 Hungerford JE, Owens GK, Argraves WS, Little CD. Development of the aortic vessel wall as defined by vascular smooth muscle and extracellular matrix markers. Dev Biol 1996; 178: 375-392 . 10.1006/dbio.1996.0225
- 12 Eriksson J, Löfberg J. Development of the hypochord and dorsal aorta in the zebrafish embryo (Danio rerio). J Morphol 2000; 244: 167-176 . 10.1002/(SICI)1097-4687(200006)244:3<167::AID-JMOR2>3.0.CO;2-J
- 13 Hunt P, Clarke JD, Buxton P, Ferretti P, Thorogood P. Stability and plasticity of neural crest patterning and branchial arch Hox code after extensive cephalic crest rotation. Dev Biol 1998; 198: 82-104 . 10.1006/dbio.1998.8886
- 14 Bockman DE, Redmond ME, Kirby ML. Alteration of early vascular development after ablation of cranial neural crest. Anat Rec 1989; 225: 209-217 . 10.1002/ar.1092250306
- 15 Bockman DE, Redmond ME, Waldo K, Davis H, Kirby ML. Effect of neural crest ablation on development of the heart and arch arteries in the chick. Am J Anat 1987; 180: 332-341 . 10.1002/aja.1001800403
- 16 Bergwerff M, DeRuiter MC, Hall S, Poelmann RE, Gittenberger-de Groot AC. Unique vascular morphology of the fourth aortic arches: possible implications for pathogenesis of type-B aortic arch interruption and anomalous right subclavian artery. Cardiovasc Res 1999; 44: 185-196 . 10.1016/S0008-6363(99)00186-8
- 17 Kirby ML. Cellular and molecular contributions of the cardiac neural crest to cardiovascular development. Trends Cardiovasc Med 1993; 3: 18-23 . 10.1016/1050-1738(93)90023-Y
- 18 Porras D, Brown CB. Temporal-spatial ablation of neural crest in the mouse results in cardiovascular defects. Dev Dyn 2008; 237: 153-162 . 10.1002/dvdy.21382
- 19 Kirby ML, Hunt P, Wallis K, Thorogood P. Abnormal patterning of the aortic arch arteries does not evoke cardiac malformations. Dev Dyn 1997; 208: 34-47 . 10.1002/(SICI)1097-0177(199701)208:1<34::AID-AJA4>3.0.CO;2-2
- 20 Yashiro K, Shiratori H, Hamada H. Haemodynamics determined by a genetic programme govern asymmetric development of the aortic arch. Nature 2007; 450: 285-288 . 10.1038/nature06254
- 21 Larsen W.. Development of the heart. In: Human Embryology. New York: Churchill Livingstone; 1997. , p. 169-187
- 22 Bruneau B. The developmental genetics of congenital heart disease. Nature 2008; 451: 943-948 . 10.1038/nature06801
- 23 Nishibatake M, Kirby ML, Van Mierop LH. Pathogenesis of persistent truncus arteriosus and dextroposed aorta in the chick embryo after neural crest ablation. Circulation 1987; 75: 255-264 . 10.1161/01.CIR.75.1.255
- 24 Hutson MR, Kirby ML. Model systems for the study of heart development and disease. Cardiac neural crest and conotruncal malformations. Semin Cell Dev Biol 2007; 18: 101-110 . 10.1016/j.semcdb.2006.12.004
- 25 Restivo A, Piacintini G, Placidi S, Saffirio C, Marino B. Cardiac outflow tract: a review of some embryogenetic aspects of the conotruncal region of the heart. Anat Rec A Discov Mol Cell Evol Biol 2006; 288: 936-943 . 10.1002/ar.a.20367
- 26 Beall AC, Rosenquist TH. Smooth muscle cells of neural crest origin form the aorticopulmonary septum in the avian embryo. Anat Rec 1990; 226: 360-366 . 10.1002/ar.1092260313
- 27 Kappetein A, Gittenberger-de Groot A, Zwinderman A, Rohmer J, Poelmann R, Huysmans H. The neural crest as a possible pathogenetic factor in coarctation of the aorta and bicuspid aortic valve. J Thorac Cardiovasc Surg 1991; 102: 830-836
- 28 Anderson RH, Webb S, Brown NA, Lamers W, Moorman A. Development of the heart: (3) formation of the ventricular outflow tracts, arterial valves, and intrapericardial arterial trunks. Heart 2003; 89: 1110-1118 . 10.1136/heart.89.9.1110
- 29 Qayyum SR, Webb S, Anderson RH, Verbeek FJ, Brown NA, Richardson MK. Septation and valvar formation in the outflow tract of the embryonic chick heart. Anat Rec 2001; 264: 273-283 . 10.1002/ar.1162
- 30 Webb S, Qayyum SR, Anderson RH, Lamers WH, Richardson MK. Septation and separation within the outflow tract of the developing heart. J Anat 2003; 202: 327-342 . 10.1046/j.1469-7580.2003.00168.x
- 31 Maron B, Hutchins G. The development of the semilunar valves in the human heart. Am J Pathol 1974; 74: 333-344
- 32 Fernández B, Durán AC, Fernández-Gallego T, Fernández MC, Such M, Arqué JM. , et al. Bicuspid aortic valves with different spatial orientations of the leaflets are distinct etiologic entities. J Am Coll Cardiol 2009; 54: 2312-2318 . 10.1016/j.jacc.2009.07.044
- 33 Cho H, Harrison K, Schwartz O, Kehrl JH. The aorta and heart differentially express RGS (regulators of G-protein signalling) proteins that selectively regulate sphingosine 1-phosphate, angiotensin II and endothelin-1 signalling. Biochem J 2003; 371: 973-980 . 10.1042/BJ20021769
- 34 Downes M, Koopman P. SOX18 and the transcriptional regulation of blood vessel development. Trends Cardiovasc Med 2001; 11: 318-324 . 10.1016/S1050-1738(01)00131-1
- 35 Yang X, Castilla LH, Xu X, Li C, Gotay J, Weinstein M. , et al. Angiogenesis defects and mesenchymal apoptosis in mice lacking SMAD5. Development 1999; 126: 1571-1580
- 36 Zhong TP, Rosenberg M, Mohideen MA, Weinstein B, Fishman MC. gridlock, an HLH gene required for assembly of the aorta in zebrafish. Science 2000; 287: 1820-1824 . 10.1126/science.287.5459.1820
- 37 Jaffe M, Sesti C, Washington IM, Du L, Dronadula N, Chin MT. , et al. Transforming growth factor β signaling in myogenic cells regulates vascular morphogenesis, differentiation and matrix synthesis. Arterioscler Thromb Vasc Biol 2012; 23: e1-e11 . 10.1161/ATVBAHA.111.238410
- 38 Topouzis S, Majesky M. Smooth muscle lineage diversity in the chick embryo: two types of aortic smooth muscle cell differ in growth and receptor-mediated transcriptional responses to transforming growth factor-β. Dev Biol 1996; 178: 430-445 . 10.1006/dbio.1996.0229
- 39 Patterson LJ, Gering M, Patient R. Scl is required for dorsal aorta as well as blood formation in zebrafish embryos. Blood 2005; 105: 3502-3511 . 10.1182/blood-2004-09-3547
- 40 Cha YI, Kim SH, Solnica-Krezel L, DuBois RN. Cyclooxygenase-1 signaling is required for vascular tube formation during development. Dev Biol 2005; 282: 274-283 . 10.1016/j.ydbio.2005.03.014
- 41 Ben Abdelkhalek H, Beckers A, Schuster-Gossler K, Pavlova MN, Burkhardt H, Lickert H. , et al. The mouse homeobox gene Not is required for caudal notochord development and affected by the truncate mutation. Genes Dev 2004; 18: 1725-1736 . 10.1101/gad.303504
- 42 Kuo CT, Morrisey EE, Anandappa R, Sigrist K, Lu MM, Parmacek MS. , et al. GATA4 transcription factor is required for ventral morphogenesis and heart tube formation. Genes Dev 1997; 11: 1048-1060 . 10.1101/gad.11.8.1048
- 43 Holzenberger M, Ayer-Le Lièvre, Robert L. Tropoelastin gene expression in the developing vascular system of the chicken: an in situ hybridization study. Anat Embryol 1993; 188: 481-492
- 44 Abdulla RI, Slott EF, Kirby ML. Proteins associated with cardiac neural crest in the pharyngeal region of early chick embryos. Pediatr Res 1993; 33: 43-47 . 10.1203/00006450-199301000-00009
- 45 Biben C, Hatzistavrou T, Harvey R. Expression of NK-2 class homeobox gene Nkx2-6 in foregut endoderm and heart. Mech Dev 1998; 73: 125-127 . 10.1016/S0925-4773(98)00037-9
- 46 Calmont A, Ivins S, Van Bueren KL, Papangeli I, Kyriakopoulou V, Andrews WD. , et al. Tbx1 controls cardiac neural crest cell migration during arch artery development by regulating Gbx2 expression in the pharyngeal ectoderm. Development 2009; 136: 3173-3183 . 10.1242/dev.028902
- 47 Gittenberger-de Groot AC, Azhar M, Molin DG. Transforming growth factor beta-SMAD2 signaling and aortic arch development. Trends Cardiovasc Med 2006; 16: 1-6 . 10.1016/j.tcm.2005.09.006
- 48 Waldo KL, Lo CW, Kirby ML. Connexin 43 expression reflects neural crest patterns during cardiovascular development. Dev Biol 1999; 208: 307-323 . 10.1006/dbio.1999.9219
- 49 Yanagisawa H, Hammer RE, Richardson JA, Williams SC, Clouthier DE, Yanagisawa M. Role of endothelin-1/endothelin-A-receptor-mediated signaling pathway in the aortic arch patterning in mice. J Clin Invest 1998; 102: 22-33 . 10.1172/JCI2698
- 50 Morishima M, Yanagisawa H, Yanagisawa M, Baldini A. Ece1 and Tbx1 define distinct pathways to aortic arch morphogenesis. Dev Dyn 2003; 228: 95-104 . 10.1002/dvdy.10358
- 51 Gavalas A, Trainor P, Ariza-McNaughton L, Krumlauf R. Synergy between Hoxa1 and Hoxb1: the relationship between arch patterning and the generation of cranial neural crest. Development 2001; 128: 3017-3027
- 52 Kameda Y. Hoxa3 and signaling molecules involved in aortic arch patterning and remodeling. Cell Tissue Res 2009; 336: 165-178 . 10.1007/s00441-009-0760-7
- 53 Liao J, Aggarwal VS, Nowotschin S, Bondarev A, Lipner S, Morrow BE. Identification of downstream genetic pathways of Tbx1 in the second heart field. Dev Biol 2008; 316: 524-537 . 10.1016/j.ydbio.2008.01.037
- 54 Molin DG, Poelmann RE, DeRuiter MC, Azhar M, Doetschman T, Gittenberger-de Groot AC. Transforming growth factor beta-SMAD2 signaling regulates aortic arch innervation and development. Circ Res 2004; 95: 1109-1117 . 10.1161/01.RES.0000150047.16909.ab
- 55 Ohnemus S, Kanzler B, Jerome-Majewska L, Papaioannou VE, Boehm T, Mallo M. Aortic arch and pharyngeal phenotype in the absence of BMP-dependent neural crest in the mouse. Mech Dev 2002; 119: 127-135 . 10.1016/S0925-4773(02)00345-3
- 56 Schäfer K, Neuhaus P, Kruse J, Braun T. The homeobox gene Lbx1 specifies a subpopulation of cardiac neural crest necessary for normal heart development. Circ Res 2002; 92: 73-80 . 10.1161/01.RES.0000050587.76563.A5
- 57 Thomas T, Kurihara H, Yamagishi H, Kurihara Y, Yazaki Y, Olson EN. , et al. A signaling cascade involving endothelin-1, dHAND and msx1 regulates development of neural-crest-derived branchial arch mesenchyme. Development 1998; 125: 3005-3014
- 58 Hutson MR, Zhang P, Stadt HA, Sato AK, Li YX, Burch J. , et al. Cardiac arterial pole alignment is sensitive to FGF8 signaling in the pharynx. Dev Biol 2006; 295: 486-497 . 10.1016/j.ydbio.2006.02.052
- 59 van Gijn M, Blankesteijn W, Smits J, Hierck B, Gittenberger-de Groot A. Frizzled 2 is transiently expressed in neural crest-containing areas during development of the heart and great arteries in the mouse. Anat Embryol 2001; 203: 185-192
- 60 Vitelli F, Zhang Z, Huynh T, Sobotka A, Mupo A, Baldini A. Fgf8 expression in the Tbx1 domain causes skeletal abnormalities and modifies the aortic arch but not the outflow tract phenotype of Tbx1 mutants. Dev Biol 2006; 295: 559-570 . 10.1016/j.ydbio.2006.03.044
- 61 Vitelli F, Taddei I, Morishima M, Meyers EN, Lindsay EA, Baldini A. A genetic link between Tbx1 and fibroblast growth factor signaling. Development 2002; 129: 4605-4611
- 62 Randall V, McCue K, Roberts C, Kyriakopoulou V, Beddow S, Barrett AN. , et al. Great vessel development requires biallelic expression of Chd7 and Tbx1 in pharyngeal ectoderm in mice. J Clin Invest 2009; 119: 3301-3310 . 10.1172/JCI37561
- 63 Scambler PJ. 22q11 deletion syndrome: a role for TBX1 in pharyngeal and cardiovascular development. Pediatr Cardiol 2010; 31: 378-390 . 10.1007/s00246-009-9613-0
- 64 Bergwerff M, Gittenberger-de Groot AC, Wisse LJ, DeRuiter MC, Wessels A, Martin JF. , et al. Loss of function of the Prx1 and Prx2 homeobox genes alters architecture of the great elastic arteries and ductus arteriosus. Virchows Arch 2000; 436: 12-19 . 10.1007/PL00008193
- 65 Farrell MJ, Stadt H, Wallis KT, Scambler P, Hixon RL, Wolfe R. , et al. HIRA, a DiGeorge syndrome candidate gene, is required for cardiac outflow tract septation. Circ Res 1999; 84: 127-135 . 10.1161/01.RES.84.2.127
- 66 Heathcote K, Braybrook C, Abushaban L, Guy M, Khetyar ME, Patton MA. , et al. Common arterial trunk associated with a homeodomain mutation of NKX2.6. Hum Mol Genet 2005; 14: 585-593 . 10.1093/hmg/ddi055
- 67 Mesbah K, Harrelson Z, Théveniau-Ruissy M, Papaioannou VE, Kelly RG. Tbx3 is required for outflow tract development. Circ Res 2008; 103: 743-750 . 10.1161/CIRCRESAHA.108.172858
- 68 Nie X, Brown CB, Wang Q, Jiao K. Inactivation of Bmp4 from the Tbx1 expression domain causes abnormal pharyngeal artery and cardiac outflow tract remodeling. Cells Tissue Organs 2011; 193: 393-403 . 10.1159/000321170
- 69 Washington Smoak I, Byrd NA, Abu-Issa R, Goddeeris MM, Anderson R, Morris J. , et al. Sonic hedgehog is required for cardiac outflow tract and neural crest cell development. Dev Biol 2005; 283: 357-372 . 10.1016/j.ydbio.2005.04.029
- 70 Kim RY, Robertson EJ, Solloway MJ. Bmp6 and Bmp7 are required for cushion formation and septation in the developing mouse heart. Dev Biol 2001; 235: 449-466 . 10.1006/dbio.2001.0284
- 71 Xu H, Morishima M, Wylie JN, Schwartz RJ, Bruneau BG, Lindsay EA. , et al. Tbx1 has a dual role in the morphogenesis of the cardiac outflow tract. Development 2004; 131: 3217-3227 . 10.1242/dev.01174
- 72 Vincentz JW, McWhirter JR, Murre C, Baldini A, Furuta Y. Fgf15 is required for proper morphogenesis of the mouse cardiac outflow tract. Genesis 2005; 41: 192-201 . 10.1002/gene.20114
- 73 Chen YH, Ishii M, Sun J, Sucov HM, Maxson Jr RE. Msx1 and Msx2 regulate survival of secondary heart field precursors and post-migratory proliferation of cardiac neural crest in the outflow tract. Dev Biol 2007; 308: 421-437 . 10.1016/j.ydbio.2007.05.037
- 74 Liao J, Aggarwal VS, Nowotschin S, Bondarev A, Lipner S, Morrow BE. Identification of downstream genetic pathways of Tbx1 in the second heart field. Dev Biol 2008; 316: 524-537 . 10.1016/j.ydbio.2008.01.037
- 75 Parisot P, Mesbah K, Théveniau-Ruissy M, Kelly RG. Tbx1, subpulmonary myocardium and conotruncal congenital heart defects. Birth Defects Res A Clin Mol Teratol 2011; 91: 477-484 . 10.1002/bdra.20803
- 76 Raid R, Krinka D, Bakhoff L, Abdelwahid E, Jokinen E, Kärner M. , et al. Lack of Gata3 results in conotruncal heart anomalies in mouse. Mech Dev 2009; 126: 80-89 . 10.1016/j.mod.2008.10.001
- 77 Jiang X, Rowitch DH, Soriano P, McMahon AP, Sucov HM. Fate of the mammalian cardiac neural crest. Development 2000; 127: 1607-1616
- 78 Hove JR, Köster RW, Forouhar AS, Acevedo-Bolton G, Fraser SE, Gharib M. Intracardiac fluid forces are an essential epigenetic factor for embryonic cardiogenesis. Nature 2003; 421: 172-177 . 10.1038/nature01282
- 79 Ewart AK, Morris CA, Ensing GJ, Loker J, Moore C, Leppert M. , et al. A human vascular disorder, supravalvular aortic stenosis, maps to chromosome 7. Proc Natl Acad Sci U S A 1993; 90: 3226-3230 . 10.1073/pnas.90.8.3226
- 80 Galvin KM, Donovan MJ, Lynch CA, Meyer RI, Paul RJ, Lorenz JN. , et al. A role for Smad6 in development and homeostasis of the cardiovascular system [Letter]. Nat Genet 2000; 24: 171-174 . 10.1038/72835
- 81 Martin LJ, Ramachandran V, Cripe LH, Hinton RB, Andelfinger G, Tabangin M. , et al. Evidence in favor of linkage to human chromosomal regions 18q, 5q and 13q for bicuspid aortic valve and associated cardiovascular malformations. Hum Genet 2007; 121: 275-284 . 10.1007/s00439-006-0316-9
- 82 Sans-Coma V, Cardo M, Durán AC, Franco D, Fernández B, Arqué JM. Evidence for quantitative genetic influence on the formation of aortic valves with 2 leaflets in the Syrian hamster. Cardiol Young 1993; 3: 132-140
- 83 Olson TM, Michels VV, Urban Z, Csiszar K, Christiano AM, Driscoll DJ. , et al. A 30kb deletion within the elastin gene results in familial supravalvular aortic stenosis. Hum Mol Genet 1995; 9: 1677-1679
- 84 Ruddy JM, Jones JA, Spinale FG, Ikonomidis JS. Regional heterogeneity within the aorta: relevance to aneurysm disease. J Thorac Cardiovasc Surg 2008; 136: 1123-1130 . 10.1016/j.jtcvs.2008.06.027
- 85 Milewicz DM, Guo DC, Tran-Fadulu V, Lafont AL, Papke CL, Inamoto S. , et al. Genetic basis of thoracic aortic aneurysms and dissections: focus on smooth muscle cell contractile dysfunction. Annu Rev Genom Hum Genet 2008; 9: 283-302 . 10.1146/annurev.genom.8.080706.092303
- 86 Wang L, Guo DC, Cao J, Gong L, Kamm KE, Regalado E. , et al. Mutations in myosin light chain kinase cause familial aortic dissections. Am J Hum Genet 2010; 87: 701-707 . 10.1016/j.ajhg.2010.10.006
- 87 The National Registry of Genetically Triggered Thoracic Aortic Aneurysms and Cardiovascular Conditions (GenTAC). Available online at: https://gentac.rti.org/Home.aspx