
Subscribe to RSS
DOI: 10.1055/a-2309-5581
A Comprehensive Review of Capsaicin and Its Role in Cancer Prevention and Treatment

Abstract
This study examines the fundamental chemical mechanisms responsible for capsaicin's advantageous impact on cancer, specifically investigating its influence on several biological processes such as inflammation in cancer metastasis, apoptosis, angiogenesis, and cellular proliferation. This entity's connections with other signaling pathways, including PI3K/AKT, NF-B, and TRPV channels, which have been linked to tumor growth, are thoroughly examined in this work. This study presents a thorough analysis of preclinical studies and clinical trials investigating the efficacy of capsaicin in treating many forms of cancer, such as breast, prostate, colorectal, pancreatic, and others. Through tests conducted in both live organisms and laboratory settings, it has been determined that capsaicin has the ability to inhibit tumor growth and induce apoptosis in cancer cells. (in vitro and in vivo). Researchers have also looked at the results of combining capsaicin with chemotherapy medications in traditional treatment. The efficacy and bioavailability of capsaicin as a viable medicinal drug are being studied, along with ways to improve its clinical value. The present investigation carefully assesses the challenges and potential options for maximizing the therapeutic benefits of capsaicin, including customized drug delivery and personalized therapeutic strategies. In finalization, this comprehensive investigation brings together the evidence currently obtainable on the anticancer properties of capsaicin, underscoring its potential as an autonomous treatment option in the struggle against cancer. Capsaicin is a compound of significant relevance for continuing research and clinical exploration in the field of cancer treatment due to its diverse mechanisms of action and ability for boosting prevailing therapy approaches.
#
Keywords
capsaicin - breast cancer - prostate cancer - colorectal cancer - pancreatic cancer - protein inhibitionIntroduction
Capsaicin (trans-8-methyl-N-vanillyl-6-zonisamide) is essential for chilli peppers to serve as an anticancer drug. The physiological impacts of capsaicin include anti-inflammatory, antioxidant, and anti-obesity characteristics. Furthermore, capsaicin has significant anticancer effects that protect against various malignancies. According to research, capsaicin promotes apoptosis in gastric malignancies by up-regulating p53 and proapoptotic proteins such as BAX, caspase-3, and caspase-8 [1] [2]. Because they target numerous signaling networks involved in the formation and progression of tumors, natural chemicals have attracted crucial interest in recent years in the pursuit of innovative cancer therapeutic strategies [3].
Capsaicin, the primary molecule in chilli peppers, is the component that gives them a uniquely smoky flavor. Assessing the detrimental effects of capsaicin (TRPV1) attributed to the exploration of transient receptor potential vanilloid subfamily member 1. Considering this specific receptor is present on essential sensory afferents, deliberate administration of capsaicin to activate just pain afferents is now being studied in human and animal models for a multitude of potential applications. Capsaicin diverges from various other naturally occurring abrasive compounds in that it generates preliminary neural stimulated activity. In the extensive refractory phase that following, the formerly activated neurons become impervious to other stressors. This de-functionalization process has been used to the benefit of capsaicin therapies for a number of painful ailments. We assessed a wide range of documents on the uses of capsaicin in healthcare and how it performs. Capsaicin's therapeutic benefits have been related to a variety of cancers, obesity, gastrointestinal and cardiovascular illnesses, neurogenic bladder, and dermatological ailments. In order to clarify with these consequences, a number of speculations have been offered. It's noteworthy the amount of time TRPV1 is essential for the vast majority of these pharmacological consequences [4].
The primary constituent in chilli peppers, which are the most common chilli in countries around the globe and a member of the Capsicum genus of plants, is called capsaicin. Capsaicin and related compounds are belonging to the class of chemical compounds that occur naturally known as capsaicinoids. Capsaicin, which gives chillies their distinctly spiciness, has been suggested to be formed by the plant as a form of natural resistance against fungi and herbivores. Researchers have been studying the ways Capsaicin impacts human beings since over a century ago. Hogyes specified the burning and hyperemia that a capsicum extract provoked when applied to human skin in 1878 [5].
Further investigations on animals revealed that intravenous administration of Capsicum extract dropped blood pressure, increased gastric and salivary secretion, and enhanced intestinal Q2 efficiency. As a result, studies aimed at using capsaicin as a form of treatment for a wide range of clinical conditions is expanding [4].
The plant genera in the botanical family Capsicum, typically referred to as chilli pepper plants, generate capsaicin, a naturally occurring alkaloid, composed of trans-8-methyl-N-vanillyl-6-zonisamide. The chemical mechanism underlying capsaicin is C18H27NO3. It is a white, crystalline powder that has minimal color or odour and is exceptionally volatile, pungent, hydrophobic, and astringent. As a prerequisite for the chilli pepper to generate vanillyl amine, a branched-chain fatty acid must be further stated [6]. In order to synthesize it in terms of revenue, vanillyl amine is synthesized by combining with the paprika-derived chloride of 7-methyloct-5-ene-1-carboxylic acid. Furthermore, dried, ripe Capsicum frutescens L. (chili pepper) fruits can also be finely pulverized in order to produce the output. Liquid formulation, liquid spray ground level, and dry powder are the three distinct kinds of authorized formulations. It has been suggested that dehydrogenation transforms capsaicin into unique macrocyclic, -diene, and -imide metabolites. It has been shown that the cytochrome P 1A1, 1A2, 2B6, 2C8, 2C9, 2C19, 2D6, and 3A4 enzymes activate the metabolism [7].
Additionally, there does not exist confirmation that capsaicin's interaction with TRVP1 receptors enhances its anticancer effects, despite the fact that its anticancer characteristics have been studied in a number of carcinogenic compounds and several theories have been put up to explain this mode of action [8]. Studies has been done to investigate how capsaicin affects human cancer cells derived from an immortal cell line that is frequently employed in empirical research. Still, a few findings indicate towards an TRPV1-mediated rise in intracellular calcium that triggers the apoptotic pathway. They highlighted effects which mean diminished cancer cell survival and proliferation, G2/M phase cell cycle slow down and loss of the mitochondrial membrane potential, which triggered caspase-9 and subsequently caused apoptosis [9].
Capsaicin-induced apoptosis in pancreatic cancer cells was demonstrated to be triggered on by impairment in mitochondrial membrane potential. Additionally, they established how capsaicin reduced the activity of the enzyme’s catalase, glutathione peroxidase, and superoxide dismutase. Furthermore, they confirmed that while having no effect on healthy cells, In cancer cells, capsaicin inhibits the chain of electron transportation networks 1 and 3. Recent research on pancreatic cancer has demonstrated that capsaicin stimulates caspase-3, induces G0/G1 phase cell arrest, and reduces the survival of cancer cells in a dose-dependent manner [10] [11]. Based on studies using prostate cancer cell lines, capsaicin inhibits the activity of nuclear factor-kappa (NF-k) and tumor necrosis factor-alpha (TNF-alpha). Numerous cancers, including hepatocellular carcinoma, breast cancer, colon cancer, gastric cancer, and leukemia, have shown comparable evidence of the benefit of therapy. Reactive oxygen species are created when complexes 1 and 3 are disrupted, and these species are fatal to cells in human beings. In the human pancreatic cancer cell line PANC-1, capsaicin-induced mortality is mediated by the phosphoinositide 3-kinase/Akt mechanism, according to recent studies on the condition, Capsaicin suppresses the activity of nuclear factor-kappa (NF-k) and tumor necrosis factor-alpha (TNF-alpha) [12], according to investigations regarding the way it affects prostate malignancy cell lines, corresponding indications of the anticancer impact has been observed in several cancer cases, such as hepatocellular carcinoma [13], colon cancer [14], gastric cancer [15], breast cancer [14] [16] [17] and leukemia [18].
#
Breast Cancer
Capsaicin Prevents HUVEC Proliferation provoked by VEGF
In order to ascertain capsaicin's in-vitro antiangiogenic activity, its inhibitory impact on VEGF-induced endothelial cell proliferation was evaluated. HUVECs were stimulated with several doses of capsaicin for 30 min before being exposed for 48 hours at 10 ng/ml of VEGF. In a dose-dependent manner, capsaicin inhibited the proliferation of HUVECs stimulated by VEGF, with a half-maximum suppression seen at 5 μM. Up to 25 μM, capsaicin exhibited no effect on the conventional proliferation of HUVECs without VEGF stimulation, suggesting that these inhibitory effects weren't brought on by the substance cytotoxicity in endothelial cells. Additionally, other kinds of investigated cells, specifically HK cells, which mimic follicular dendritic cells, and RAW 264.7 were unable to demonstrate any cytotoxic impact associated with the chemical up to 50 μM. Using the 3[H]thymidine incorporation assay, the impact of capsaicin on the DNA synthesis of HUVECs was investigated. VEGF (10 ng/ml) substantially boosted the efficiency of DNA synthesis in HUVECs, and capsaicin completely neutralized this effect.
#
Capsaicin Triggers Endothelial Cells to Arrest in G1 via Down-Regulating Cyclin D1
Examining how capsaicin influences the course of the cell stage using a fluorescence-activated cell sorter. Following a 20-hour incubation period with or without 10 ng/ml VEGF, we assessed the percentage of HUVECs in the G0/G1, S, and G2-M phases of the cell cycle. Due to VEGF, HUVECs reached the S phase. However, a dose-dependent decrease in S phase entrance was seen when capsaicin was administered. At 5 M capsaicin, the effect's half-maximum was noted. Our findings thus suggest that capsaicin influences the G1 to S phase of cell development. Cyclin D1 facilitates the transition of cells from the G1 to the S phase. The passage of cells from the G1 to the S phase is aided by cyclin D1. The control of cyclin D1 was examined as a potential mechanism by which capsaicin mediates its antiproliferative actions. Protein kinase triggered by cyclins and p125, the two downward KDR/Flk-1 signaling molecules, showed up more frequently when HUVECs had been subjected to VEGF for different periods of latency. Even at a 25 M dose, capsaicin had no effect on the autophosphorylation of KDR/Flk-1 caused by VEGF. Fortunately, capsaicin drastically and dose-dependently suppressed VEGF-induced activation in both p38 mitogen-activated protein kinase FAK and p125. The activation of kinase controlled by extracellular signals imposed by VEGF wasn't impacted by capsaicin (up to 25 M). It has been demonstrated that VEGF induces endothelial cell migration, proliferation, and tube formation, and it has been shown that this is entirely reliant on the AKT/NO pathway. AKT (Ser-473) and endothelial NO synthase (eNOS; Ser-1177) phosphorylation are triggered by VEGF, and both of these mechanisms are vital for VEGF-induced angiogenic growth. Therefore, utilizing primers against the phosphorylated forms of eNOS (Ser-1177) and AKT (Ser-473), the effects of capsaicin on VEGF-induced phosphorylation of these two proteins were assessed. Capsaicin attenuated VEGF-induced phosphorylation of AKT and eNOS in a dose-dependent way, with an impact at half its peak at 5 M. According to the previously cited research, capsaicin can prevent endothelial cells' amplification of the VEGF-induced AKT/eNOS signaling pathways [19] [20] [21].
In In-Vivo studies, Angiogenesis caused by tumor is blocked by capsaicin. Using the chick chorioallantoic membrane experiment, it was then determined if capsaicin is effective in preventing tumor-induced angiogenesis. Collagen gels type I induced evident blood arteries oozing into collagen production sponges. when HT1080 cells were enclosed in them as opposed to medium separately. While having no adverse consequences on pre-existing blood vessels, capsaicin reduced tumor-induced angiogenesis. While comparing to normal media alone, cancer cells significantly (2.8 times) improved the number of newly formed blood vessels [22].
#
Effectiveness of capsaicin on breast cancer cell apoptosis, propagation, and cell phase
Using the MTT (3-(4, 5-dimethylthiazol- 2-yl)-2, 5-diphenyltetrazolium bromide) assay, the ability of capsaicin to inhibit the proliferation of a variety of ER-positive and -negative carcinoma cell lines was investigated. All the examined breast carcinoma cell lines (MCF-7, SKBR-3, MDA-MB231, T47D, and BT- 474) depends on dosage decline in proliferative activity in response to capsaicin. Assessing ER-negative SKBR-3 cells are additional cell lines for breast cancer that were examined, it emerged that ER-negative MDA-MB231 cells were less susceptible for substance. Treatment with capsaicin had no discernible effect on the growth of the un-transformed human lactating epithelial cell line MCF-10A. BT-474, SKBR-3, and MDA-MB231 cells had their cell-cycle segregation examined using the technique of flow cytometry. Capsaicin treatment impacted all cell lines, resulting in a dose- and time-varying slowdown in cell growth during the G0/G1 phase and a reduction in S phase cells. Following a 72-hour treatment period, propidium iodide (PI) and Annexin V-FITC (fluorescein isothiocyanate) were used to dye BT-474, SKBR-3, MDA-MB231, and MCF-7 cells in order to evaluate the ability of capsaicin to induce apoptosis. After 24 hours of capsaicin treatment, the percentage of apoptotic cells increased dramatically in all four breast cancer cell lines, and by 72 hours, over 80% of the cells were positive for Annexin V-FITC and PI. Less than 15% of cells exposed to the diluent control (dimethyl sulfoxide, DMSO) exhibited positive after 72 hours.
Capsaicin manipulates the EGFR/HER-2 pathway. Breast cancer develops partly because of large part of the actions of EGFR and HER-2, which can cause tumor development and invasion by signalling through intracellular kinases. In ER-positive BT-474, ER-negative SKBR-3, and MDA-MB231 lineages of cells, capsaicin was examined for its potential to reduce the levels of EGFR proteins associated to paths. In BT-474 and SKBR-3 cells, capsaicin significantly lowered the expression of the proteins EGFR and HER-2. In contrast, following 24 h of therapy, the medication minimally reduced EGFR and HER-2 levels in MDA-MB231 cells. The mRNA level did not demonstrate the modulatory effect of capsaicin on the EGFR and HER-2 expression. Furthermore, after being subjected to capsaicin, the cell lines BT-474, SKBR-3, and MDA-MB231 exhibited reduced levels of the cell-phase regulator cyclin D1 and an increase in the cyclin-dependent kinase inhibitor p27KIP1. This was associated with caspase-3, -7, and -9 stimulation and the discovery of PARP (poly-ADP ribose polymerase) pectoral items that are distinctive to apoptosis. Cyclin A, B1, cdk-2, and p21WAF1 expression levels were unaffected in capsaicin-treated cell varieties.
Despite serum-starved cells' stimulation with EGF, capsaicin reduced the enzyme activity of extracellular-regulated kinase (ERK), a crucial downstream signalling molecule in the EGFR/HER-2 signalling cascade. Even in the recognized intrinsically active ERK-containing MDA-MB231 cells, capsaicin suppressed the levels of phosphorylated ERK. In contrast, capsaicin had no effect on the levels of activated AKT or signal transducer and activator of transcription 3 (STAT3) in any of the previously stated cell lines.
#
Effect of capsaicin on breast tumor cells EGFR-dependent mobility
Breast cancer departure may be influenced by EGFR/HER-2 overexpression and the next-generation transmitting molecule ERK [23] [24]. Baselga, J investigated Capsaicin's ability to prevent breast cancer cells from migrating in an EGF-dependent manner using a Boyden chamber experiment. The bottom chamber's EGF could be approached by oversight cells from the breast cancer cell lines BT-474, SKBR-3, and MDA-MB231. Conversely, when the investigation had been carried out with capsaicin present, cell migration slowed down in all cell lines [23].
#
Effect of capsaicin on the development of lumps in the breast triggered by DMBA
Using 30 BALB/c mice's breast glands, growth hormones were administered either with Capsaicin or a diluent control, and the glands were subsequently subjected to either 7,12-dimethyl-benz[a]anthracene (DMBA) for 24 hours to evaluate if Capsaicin was beneficial for suppressing the occurrence of alveolar and ductal lesions. After 14 days of DMBA treatment, the glands continued to swell. During the DMBA treatment, capsaicin eliminated breast ductal and alveolar lesions. Biopsy studies showed that even when healthy tissue of the mammary fat cells and mammary ducts was preserved, under-treatment with capsaicin decreased malignant alveolar and secretory lesions. The glands treated with capsaicin at the level employed in the organ culture experiments did not exhibit any histological evidence of damage. The percentage decrease in lesion formation was calculated by comparing the percentage of DMBA-induced lesions in the groups treated with capsaicin and diluent control. According to that capsaicin dose-dependently blocks glands with lesions [25]. Apoptosis is promoted by capsaicin by means of AMPKa-mTOR mechanism. It was possible to ascertain that capsaicin-treated cells possessed vacuoles of varied sizes that housed cellular organelles by the use of microscopes with electrons. It is possible that these cavities were auto-lysosomes or auto-phagosomes. Cells were treated for 1 hour with the lysosomal inhibitors E64d/pepstatin and the inhibitor of autophagosome-lysosome fusion bafilomycin A1 (BaF1) to enhance LC3II accumulation. This procedure was utilized to show that capsaicin transforms LC3 to LC3II. Additional factors influencing the induction of autophagy included Atg5 activation, LC3 conversion, and a dose-dependent drop in p62. An investigation into AMP-dependent protein kinase (AMPKa) and mTOR was conducted because capsaicin stimulated Akt phosphorylation in MCF-7 cells (mTOR). Finally, phosphop70S6K was downregulated following AMPKa phosphorylation and mTOR dephosphorylation [Fig. 1]. For the purpose of investigating the autophagy inhibitor 3-MA for two hours prior to MCF-7 culture pretreatment in order to examine the purpose of capsaicin-induced autophagy, subjected to continuous Following 24- as well as 48-hour capsaicin treatment, the samples were then treated with Hoechst 33342 and propidium iodide (PI). The percentage of apoptotic cells in untreated cells was 0.45%; however, after being exposed for 24 and 48 hours, the percentage rose to 5.09% and 11.6%, respectively. Over the course of 24 and 48 hours, respectively, the rate of apoptosis increased to 10.4% and 21.7% in cells treated with capsaicin plus 3-MA. The results of this study show that capsaicin affects cell viability by inhibiting apoptosis and increasing autophagy through the AMPKa-mTOR regulatory mechanism [26].

#
PARP-1 is activated and deactivated by capsaicin
When DNA strands break, PARP-1 is activated. Depending on the severity of the DNA damage, PARP-1 either repairs the DNA or causes cell death. After 30 minutes of capsaicin stimulation, PARP-1 levels increased, but they quickly decreased after 12 hours. Finding the 29-kDa PARP-1, which is equivalent to its active form produced by caspase-7 splitting, took a whole day, unfortunately. Consequently, research on PARP-1's function in DNA repair was done. When membranes lacking PARP-1 were re-probed using an anti-poly (ADP-ribose) (PAR) antibody, the number of molecules of PAR enhanced with time, demonstrating that PARylated PARP-1 was not detectable. PARP-1 activation and cleavage were demonstrated in a dose-dependent trial [27] [28]. It was established that the decrease in 116-kDa PARP-1 was due to PARylation using the PARP-3AB inhibition, which completely inhibited PAR the synthesis process. Additionally, 3-AB-treated cells exhibited slightly greater levels of 116-kDa PARP-1 with greater quantities of the 29-kDa form compared to cells treated with capsaicin, which ultimately aggravated cell death. However, the cytoplasmic vacuolization that Capsaicin induced was not impacted by 3-AB. According to the findings, Cell protection is aided by capsaicin-induced PARP-1 activation [26]. Here, [Fig. 2] depicted the metabolic pathway of breast cancer metastasis previously described.

#
Colon Cancer
In each group, there were three wells. Viability testing was carried out to identify the appropriate dosages after the plate containing the cells had been incubated for two hours at 37 °C in an incubator with 5% CO2. When the AgNOR approach was used to expose Caco-2 cells to the ideal dose of capsaicin, the anticancer impact of the chilli was seen. Each group, which had been split into 4, utilised the AgNOR staining methodology.
Capsaicin potentially minimises the proliferation of cancer cells. The in-vitro investigation looks at how capsaicin affects human colon cancer colo 205 cells following exposure. The findings showed that the cytotoxic effects of capsaicin in colo 205 cells raised Ca2 and reactive oxygen species (ROS) while decreasing the amount of mitochondrial membrane potential (m) in time- and dose-dependent manners. Western blotting research revealed that increased Fas, cytochrome c, and caspase levels led to cell death. Capsaicin lowered levels of anti-apoptotic proteins like Bcl-2 while raising levels of pro-apoptotic proteins like Bax. Another way that capsaicin elicited apoptosis in colo 205 cells was by engaging caspase-8, -9, and -3 [16]
Effects of Capsaicin on Colo 205 Cell Viability detected by Flow cytometry. The proportion of viable cells decreased when the incubation period and capsaicin dosage were increased. At 500 μM, capsaicin dramatically reduced the number of viable cells by about 97%. After 24 hours of incubation and treatment at 150 μM, 50% of the viable cells were found.
According to Capsaicin Effects on Colo 205 Cell Cycle Restriction and Apoptosis, while colo 205 cells were subjected to different dosages of the chilli pepper for 24 hours of incubation, the fraction of cells in G0/G1 grew in a dose-dependent path. Apoptosis has been suggested by the cell cycle's sub-G1 groups, which were detected. The greatest proportion of apoptosis detected in all of the dosages examined in colo 205 cells occurred when the capsaicin dose elevated to 300 M, which is approximately 60%.
Impact of capsaicin on mitochondrial membrane potential (ΔΨm), ROS, and Ca2 in Colo 205 cells. When the outermost layer of mitochondrial membrane integrity is compromised and cytochrome c is liberated from the mitochondria into the cytoplasm, the cells are prompted to undergo cell death. Damage to the mitochondria is a result of ROS production, and additional ROS may be released into the cytoplasm as a result. In order to assess whether mitochondrial involvement in Capsaicin-induced apoptosis could be established, Colo 205 cells were exposed to 150 μM Capsaicin for the durations specified. Assessment of the changes in ROS production and ΔΨm revealed that the Capsaicin therapy disrupted ΔΨm. Capsaicin has been assessed for its impact on caspase-3, -8, and -9 activities as well as the concentration of live colo 205 cells utilising flow cytometric analysis and the caspase activity assay are shown in [Fig. 3]. The results are provided. After a 24-hour the incubation process capsaicin (150 M) increased caspase-3, -8, and -9 activities; however, prior to treatment, cells with caspase-3, -8, and -9 inhibitors, which led to reductions in caspase-3, -8, and -9 activities and increases in the percentage of viable cells. These results demonstrate that caspase-8, -9, and -3 activation is a key facilitator of capsaicin-induced apoptosis [29].

#
Lung Cancer
A transcription factor for genes known as hypoxia-induced component when the tumor microenvironment is hypoxic, HIF-1 regulates the expression of several genes [30] [31] (cancer and 2003,). In normoxia, the oxygen-dependent proteasomal system efficiently degrades HIF-1, but in hypoxia, it accumulates, triggering transcription of its target genes implicated in drug resistance, blood vessel development, growth, and metastasis. HIF-1 controls the expression of several enzymes associated with the metabolism of glucose, which has a role in cancer metabolism [32].
Capsaicin promotes inhibiting development of cells and activating HIF in lung cancer cells. The effects of Capsaicin on cell survival and HIF activation were explored to find the anticancer effectiveness of Capsaicin against lung cancer cells. H1299, H23, A549, and H2009 cells all experienced a considerable growth slowdown after exposure to Capsaicin, with IC50 values of 62.9, 54.5, 71.3, and 75.4 μM, respectively. Additionally, in lung cancer cells expressing the HRE-luciferase sensor gene, the impact of capsaicin on HIF-mediated transcriptional activation was studied. Capsaicin dramatically and concentration-dependently reduced HIF activation in hypoxic conditions. The findings show that capsaicin possesses anticancer benefits by preventing lung cancer cell proliferation and HIF activation.
The buildup of HIF-1 protein and the expression of the genes that it targets are both inhibited by capsaicin. We evaluated the impact of capsaicin on the concentration of HIF-1 protein in order to comprehend the fundamental mechanism of capsaicin-mediated inhibition of HIF activation. Compared to normoxic conditions, lung cancer cell lines showed a significant rise in HIF-1 protein after 6 hours of incubation under hypoxia conditions. In lung cancer cell lines, capsaicin treatment reduced the concentration-dependent accumulation of HIF-1 protein caused by hypoxia. Furthermore, in the H1299 cells employed in this investigation, capsaicin demonstrated the usual effects of capsaicin on pulmonary cancer cell lines by lowering the expression of the HIF-1 target genes glucose transporter 1 (GLUT1) and pyruvate dehydrogenase kinase 1 (PDK1).
Capsaicin lowers metabolism in the mitochondria, which prevents the generation of ATP in the cell's mitochondria shown in [Fig. 4]. Researchers also examined the effect of capsaicin on lung cancer cells' mitochondrial respiration in order to comprehend the mechanism underlying the therapy's augmentation of intracellular oxygen levels. Utilizing an XFe24 analyzer to measure OCR, the findings showed that treatment with capsaicin dramatically reduced intracellular OCR, which in turn prevented a variety of lung cancer cells from producing mitochondrial ATP. Furthermore, when capsaicin was given directly to H1299 cells, intracellular OCR and mitochondrial ATP synthesis were immediately decreased, indicating that capsaicin selectively inhibits mitochondrial respiration and reduces mitochondrial ATP synthesis. Our data show that capsaicin reduces the production of ATP and mitochondrial respiration, which limits cell growth and causes HIF-1 to drop [33].

#
Prostate Cancer
Capsaicin prevents prostate cells from growing. Initially, researchers investigated whether capsaicin had an effect on the LNCaP, PC3, and DU-145 prostate cancer cell models' propensity to proliferate. Capsaicin was less effective in reducing cell viability in DU-145 cells than in the other two cell lines, even at greater doses of 80 M and 160 M. DU-145 cells exhibited a greater degree of capsaicin sensitivity throughout necrosis. Capsaicin appears to be less effective in DU-145 cells, even though it caused apoptotic in 27% of LNCaP cells, 18% of PC3 cells, and only 9.6% of DU-145 cells.
In prostate cells, the activation of AMPK produced by capsaicin has not been mediated by CaMKK2. The upstream kinase CaMKK2 has the ability to activate AMPK. A pharmacological CaMKK2 inhibitor called STO-609 was utilized to investigate the role that this kinase played in the capsaicin-induced AMPK activation. STO-609 is cell-permeable and selective. In LNCaP and PC3 cells, pretreatment with STO-609 was unable to stop the phosphorylation of ACC or AMPK brought on by capsaicin. On the other hand, STO-609 stopped ACC or AMPK from being phosphorylated in the LKB1null cell line DU-145. These findings imply that CaMKK2 is not involved in the mechanism by which capsaicin activates AMPK in prostate cancer cells. However, in DU-145 cells devoid of LKB1, CaMKK2 could be implicated in AMPK basal phosphorylation. For AMPK and LKB1 to be activated, TRPV1 must exist. Transient Receptor Potential Vanilloid 1 (TRPV1) is a cation channel that is expressed in sensory neurons and various organs, including the prostate and prostate cancer cells. Capsaicin efficiently activates TRPV1. According to a recent study, TRPV1 is necessary for the development and carcinogenesis of cancer. However, TRPV1 agonists may work independently of receptors or via receptors to have beneficial effects on cancer. The developers reduced the expression of the TRPV1 channel by infecting cells with lentiviral viruses producing short hairpin RNA in an effort to learn more about the processes underlying AMPK activation and capsaicin-induced anticancer effects (shRNA). To generate viruses in HEK293T cells, the optimal TRPV1 sequence was cloned into the lentiviral vector (pLKO.1) [34].
In order to infect control cells, viruses with empty vectors (pLKO.1 EV) were used. By contrasting the results of the Western blotting tests with those of the negative control, the knockdown effectiveness of the TRPV1-specific shRNA was confirmed at the protein level. LKB1 phosphorylation was reduced in LNCaP and PC3 cells with TRPV1 genetic suppression, suggesting a relationship between TRPV1 and LKB1 in prostate cells. TRPV1-downregulated cells also prevented the capsaicin-induced increase in AMPK phosphorylation. The genetic ablation of TRPV1 had no effect on the capacity of DU-145 cells to activate AMPK in the presence of capsaicin because these cells do not express LKB1. The effect of escalating capsaicin doses on the clonogenic growth of prostate cancer cell lines LNCaP, PC-3, and DU-145 on soft agar was investigated to determine its impact on clonal proliferation and cell cycle. Each clonal progression was inhibited by capsaicin in a dose-dependent manner. At 5×10–4 mol/L, the growth of every cell type halted. For the purpose of investigating the cell cycle, PC-3 cells were exposed to elevated concentrations of Capsaicin or vehicle for 24 hours using flow cytometry. The population of cells in the G0/G1 phase grew in a dose-dependent manner, but the population of cells in the S phase diminished.
Apoptosis occurs in prostate cancer cells when they are exposed to capsaicin. Cells were exposed to increasing concentrations of capsaicin for 24 hours; apoptosis was assessed using the TUNEL assay. For PC-3 cells, the fraction of apoptotic cells altered in response to capsaicin from 3% at 1×10–4 mol/L to 75% at 5×10–4 mol/L, and from 9% at 1×10–4 mol/L to 93% at 5×10–4 mol/L for LNCaP cells.
The quantities of proteins related to apoptosis and the cell cycle are altered by capsaicin in a time-dependent way. To explain the molecular mechanism of Capsaicin-induced arrest of the cell cycle and apoptosis, researchers evaluated the levels of several apoptotic and cell cycle-related proteins in LNCaP cells that had been exposed to the compound at a concentration of 2×10–4 mol/L. The levels of the proteins Bax and p53 grew along with the expression of the p21 Waf1 protein after being exposed to capsaicin for 3, 6, and 12 hours.
Capsaicin's effect on the transcriptional activity of androgen-responsive reporter constructs. PSA is an androgen-responsive gene that is produced by prostate epithelial cells. Androgen-sensitive LNCaP cells were transfected with a PSA promoter/enhancer luciferase reporter vector (PSA P/E-Luc) in order to examine the impact of capsaicin on PSA transcription. For the 24-hour culture of these cells in 10% charcoal-striped FBS RPMI 1640, the options were dihydrotestosterone (1×10–8 mol/L), capsaicin (2×10–4 mol/L), or diluent control. The fold induction was determined by luciferase activity seen in control cells.
When compared to control LNCaP cells, dihydrotestosterone enhanced reporter activity by a factor of>60. Capsaicin absolutely stopped it from working. These results showed that capsaicin inhibited testosterone's ability to trans-activate the PSA enhancer/promoter. The luciferase-reporter architecture has been used in several research utilizing either ARE4-E4Lux (multimerized four consensus AREs from the PSA promoter) or a 496-bp PSA enhancer section composed of six AREs. Dihydrotestosterone (1×10–8 mol/L) raised luciferase activity for each scheme, and capsaicin subsequently stopped this kind of stimulation [12]
#
Pancreatic cancer
The impact of capsaicin on human pancreatic cancer cells' growth and activation of cell death as compared to normal acinar cells. Scientists first examined how capsaicin affected the proliferation of human pancreatic cancer cells AsPC-1 and BxPC-3 using the Sulforhodamine-B assay. At an IC50 of roughly 150 μM, capsaicin significantly and dose-dependently reduced the growth of both cell lines. High concentrations of capsaicin killed cancer cells rapidly, but they had little influence on normal acinar cells' ability to survive. According to earlier research, TRPV1 intracellular binding mediates the physiological effects of capsaicin on sensory neurons (vanilloid receptor). By first treating the cells for one hour with the TRPV1-specific antagonist 10 μM capsaicin and then treating them for twenty-four hours with 150 μM Capsaicin, it has been determined that the antiproliferative impact of capsaicin is caused by its binding to TRPV1. capsaicin inability to counteract capsaicin's cytotoxic effects suggests that TRPV1 is not the mechanism responsible for the compound's growth-suppressive actions. A cell-death experiment was utilized to ascertain the mechanism behind the anti-proliferative activities of capsaicin. A dose-dependent increase in apoptotic cells was seen in AsPC-1 and BxPC-3 cells following a 24-hour exposure to various doses of capsaicin. These multiplicities over the control cells were 1.6–3.6 and 4.5–6.0 times, in that order. DAPI labeling has added to the evidence that capsaicin caused AsPC-1 cells to undergo apoptosis. Capsaicin-treated AsPC-1 cells showed fractured or condensed nuclei. In contrast to expectations, capsaicin did not cause any apoptosis in the healthy acinar cells28.
After being exposed to capsaicin, AsPC-1 and BxPC-3 cells generate ROS: Reactive oxygen species (ROS) production has been postulated in a number of studies as a potential mechanism by which several anticancer medications induce death. We investigated the theory that ROS are produced as a consequence of capsaicin-induced apoptosis in our model. The cells were labeled with DCFDA, and the intracellular ROS production in the control and capsaicin-treated cells was assessed by flow cytometry. Fluorescent DCF is created when peroxides cleave and oxidize DCFDA. Following a 24-hour course of capsaicin administration, DCF fluorescence (the formation of ROS) improved in a dose-dependent manner. In AsPC-1 and BxPC-3 cells, for instance, DCF fluorescence increased by 1.4–1.7 and 1.3–2.1 times, respectively, in response to capsaicin administration in comparison to DMSO-treated control cells29.In a time-dependent experiment, capsaicin increased DCF fluorescence by approximately 71% relative to the control after one hour of treatment. H2O2 was also quantified in AsPC-1 cells after 150 M capsaicin administration because the peak of ROS generation occurred one hour after treatment. The H2O2concentration of capsaicin-treated cells was 23% greater than that of the control group. In contrast, normal acinar cells' DCF fluorescence did not increase in response to 300 μM of capsaicin administration. Effect of capsaicin on the potential of the mitochondrial membrane in AsPC-1 and BxPC-3 cells: Overproduction of ROS within cells frequently modifies the potential of the mitochondrial membrane. As a result, by identifying the cells containing the cationic lipophilic dye JC-1, which aggregates in the mitochondria of healthy cells in a potential-dependent manner, it was possible to ascertain the impact of capsaicin treatment on mitochondrial membrane potential. When compared to the control, capsaicin treatment dramatically reduced the mitochondrial membrane potential in the AsPC-1 (25–89%) and BxPC-3 (22–75%) cells. Capsaicin, however, had no effect on the mitochondrial membrane's potential function in normal acinar cells.
In AsPC-1 cells, capsaicin treatment causes the release of cytochrome c and the activation of the caspase-3 cascade. The aim of this study was to determine whether capsaicin therapy affects the expression of critical signaling molecules in the mitochondrial death cascade, since capsaicin modulates the potential of the mitochondrial membrane. Compared to control values after actin correction, the effects of a 24-hour treatment with 150 μM capsaicin on AsPC-1 cells were measured as a 4.2-, 9.0-, and 5.6-fold elevation in the cytosolic protein expression of Bax, cytochrome c, and AIF, respectively. Conversely, the creators noticed a 60% down-regulation in the expression of Survivin, a novel inhibitor of apoptosis (IAP) that inhibits caspase, which was just discovered. Bcl-2 protein expression was also markedly reduced as a result of capsaicin treatment28. Furthermore, the cleavage of caspase-9, caspase-3, and PARP indicates a significant activation of these enzymes resulting from the use of capsaicin treatment. There was additional proof that cytochrome c was released from pure mitochondrial and cytosolic fractions from both normal and capsaicin-treated AsPC-1 cells. Cytochrome C is transferred from the mitochondria to the cytoplasm upon capsaicin treatment. Following a 24-hour course of capsaicin therapy, BxPC3 cells demonstrated similar results. Nevertheless, neither the activation of caspase-3 nor the expression of these signaling proteins changed in the healthy acinar cells after capsaicin treatment.
Using immuno-fluorescence confocal imaging, the impact of capsaicin on the subcellular distribution of Bax and cytochrome c was further investigated. The green staining of control AsPC-1 cells indicates that Bax is primarily found in the cytoplasm. Bax is present in the mitochondria of cells that have been exposed to capsaicin, as evidenced by the orange staining of the mitochondria caused by the union of the red fluorescence of the mitochondria and the green fluorescence of Bax, which suggests that Bax has been transferred from the cytosol to the mitochondria. The fusion of cytochrome-green C's fluorescence with the red fluorescence of the staining agent Mito Tracker results in the orange staining of the mitochondria. Contrarily, Red showed that the majority of cytochrome-c was found in the control cells' mitochondria. When cells are exposed to capsaicin, the release of cytochrome c into the cytoplasm is visible as sporadic green staining outside of the red mitochondria. These results collectively imply that exposure to capsaicin triggers the intrinsic mitochondrial death pathway28 by releasing cytochrome c into the cytoplasm are shown in [Fig. 5].

#
Bladder Cancer
Evaluation of Mitochondrial Membrane Potential and ROS Production: TRPV1 activation dramatically raised ROS production in T24 cells. In comparison to the control, 100 and 200 mM of CAP increased ROS production by 51.0% and 95.2%, respectively. But in cells treated with CAP&CPZ or 100 mM of CAP in a Ca2+-free solution, ROS production did not change appreciably. When compared to the control, CAP at 100, 150, and 250 mM decreased the relative mitochondrial membrane potential (or red/green fluorescence ratio) in T24 cells after any type of stimulation by 19.5%, 43.7%, and 62.8%, respectively. Pre-treating cells with CPZ30 did not significantly alter the potential of the mitochondrial membrane.
Xenograft Growth of T24 Bladder Cancer Is Significant Accelerated by CAP: The average in vivo tumor diameter was 140.32 mm3 four weeks following transplantation. Following four weeks of treatment, which was repeated every three days with either CAP or vehicle, the tumor volumes (n=8, P=.01) were 567.02, 43.94, and 661.80, 62.03 mm3. At the conclusion of the trial, the mean body weight, hair coats, and overall activity of the two groups were similar, suggesting that CAP had little to no negative effect on those mice30. DCs derived from human monocytes are activated by CPS: Luckily, the researchers looked at TRPV1 expression and CPS action on monocyte-derived CD1a+iDCs before examining the impact of CPS-mediated cancer cell death on DC activation, in line with the inconsistent findings in the literature regarding CPS's ability to directly affect DC activation. Researchers found that TRPV1 was expressed at the protein level by CD1a+iDCs. Furthermore, viable DCs exhibited a dose-dependent increase of the CD86 costimulatory molecule and a slight but significant induction of the CD83+maturation marker following a 24-hour exposure to CPS at higher concentrations (from 10 to 250 μM) at 37°C. Furthermore, the addition of CPZ and TRPV1 antagonist marginally reduced the increase of CD86 and CD83 caused by CPS.
These results demonstrate that CPS is directly felt by immunogenic cell death (iCDs), and that this sensitivity is partially dependent on TRPV1. Furthermore, we found that the cytotoxic effect of CPS on DCs was dose-dependent, as when iCDs were treated with 150 and 250 μM CPS, respectively, which are the concentrations usually required to cause apoptosis in a variety of cancer cells, we found 25% and 50% of dead cells. To investigate whether cancer cell apoptosis mediated by CPS can result in DC activation, T24 and SD48 human bladder cancer cell lines were subjected to apoptosis. It's interesting to note that, although to a lesser extent, early-apoptotic cancer cells like T24 and SD48 cells that were exposed to CPS for 30 hours and 12 hours, respectively, also induced DC activation. According to our findings, capsazepine (CPS) activates DCs30 and causes cancer cells to die.
CD91 silencing prevents DC activation: In order to confirm that DC activation is caused by DAMPs of ICD released by CPS-mediated dying cancer cells, researchers investigated the possible role of CD91 (a shared receptor for CRT and HSP90/70). In order to achieve this, we used CD91siRNA to silence CD91 and decreased CD91 expression in CD1a+iDCs. Western blotting on iDC lysates and immune-fluorescence on the iDC surface were used to assess the silencing effectiveness after 24 hours. We found that transfection of iDC with CD91siRNA significantly decreased absolute and cell surface CD91 protein levels, in contrast to siCONTROL. When compared to cells transfected with control, an examination of the capacity of CPS-treated T24 cells to activate on siRNA-transfected iDCs revealed that both CD86 and CD83 markers were decreased whenever CD91 was targeted. These findings demonstrate that DC activation 30 is inhibited by CD91 silencing.
#
Liver Cancer
Impact of capsaicin on the structure and function of the liver: The AST, ALT, ALP, and bilirubin levels in the control group were all within normal ranges. But there were clear distinctions between the cyclophosphamide (CPM) group, the standard control, and the capsaicin (CAP)20 group. Rats were used to test the effects of capsaicin pre-treatment on body weight, liver weight, and their relationship to cyclophosphamide intoxication. The livers of rats given cyclophosphamide treatment were noticeably bigger than those of the healthy control group. When the ratio of liver weight to body weight increased, capsaicin increased by 4.80%. The liver weight to body weight ratio was significantly reduced by pretreatment with CPS by 2.86%. Rats that receive CPS have lower body weights, liver weights, and liver weight ratios31.
Capsaicin's effect on oxidative stress: Rats in the CPM group showed higher levels of hepatic lipid peroxidation (MDA) and lower levels of glutathione (GSH). Moreover, the CPS10+CPM200 and CPS20+CPM200 treatment groups demonstrated an efficacious therapeutic nature against liver damage induced by CPM. Following the administration of CPS10 and CPS20, it was observed that the GSH content increased and that the MDA level decreased in comparison to the CPM-induced group. Following CPM injection, the activity of several antioxidant enzymes-especially catalase and superoxide dismutase—was reduced; however, following both doses of CPS, the activity of these enzymes increased. There were no discernible differences between CPS20 and the control group31.
Capsaicin's effects on inflammatory cytokines and apoptotic markers: When it came to inflammatory cytokines, the higher dose of capsaicin was both more significant and advantageous than lower doses. When compared to the healthy control group, the liver of the CPM-administered group exhibited a significant increase in IL-1. Following CPS therapy, IL-1 levels decreased, and both CPS dosages were more significant and productive. TNF and other pro-inflammatory cytokines were significantly elevated during CPM therapy in comparison to the normal controls. TNF was considerably reduced by oral supplements containing CPS in the treatment groups when compared to the group that received CPM. When compared to the usual controls, CPM therapy significantly increased apoptotic indicators such as caspase-3. However, following oral supplementation with CPS (10 and 20 mg), both of the CPS-treated groups exhibited significantly lower levels of caspase-3 in comparison to the groups that received CPM. Higher dosages of CPS proved to be more significant and effective than lower doses. There are no discernible differences between the highest level of CPS concentration and the standard control31.
Capsaicin's effects on liver histopathology: After CPM therapy, there were concentrated areas with severe necrosis, inflammation, degeneration, and coagulation. It was determined that the injury score (score of 4) was incredibly high. The liver's histological composition greatly improved following pre-treatment with the lowest dose of CPS10, and the degree of liver damage was comparatively mild (score=2). The histological structure was significantly improved after pre-treatment with the maximum dose (CPS20), and the lowest damage score was (score=1). In contrast, the H&E-stained liver segment from the healthy control showed a normal hepatocyte arrangement with no changes and a destruction score of zero.
Capsaicin prevents DEN-induced liver cancer in rats: The most widely used experimental model for studying hepato-carcinogenesis is the rat HCC model developed by DEN. The rat liver cancer model induced by DEN was treated with capsaicin-loaded liposomes in order to study the biological effect of capsaicin on hepato-carcinogenesis in vivo. A control group (DEN+) was also established, but this group did not receive any treatment are shown in [Fig. 6]. Because liposomal capsaicin has a high potential for irritation, researchers have also administered it by subcutaneous splenic transposition. After 12–14 weeks of DEN treatment, the injection of liposomal capsaicin significantly reduced the risk of hepatocarcinogenesis (early stage). The corresponding counts of the surface nodules are shown in conjunction with sample images of the excised livers at 12 weeks. When compared to the control group (DEN+), the liver treated with capsaicin showed fewer tumor nodules and a smoother surface. Hematoxylin and eosin (H&E) staining revealed that the DEN+Capsaicin group also displayed fewer indicators of liver damage, such as oedema, inflammatory cell infiltration, and necrosis. The results show that capsaicin significantly affects the carcinogenesis of rat liver32.

During carcinogenesis, capsaicin inhibits HPC function and causes it to die: Researchers used immunohistochemistry analysis to determine how capsaicin affected the growth of the DEN-induced liver tumor and changed the state of HPCs. Capsaicin's exceptional ability to inhibit HPC activation was demonstrated by the significantly lower levels of EpCAM, SOX9, OV6, and the proliferation marker PCNA in the Capsaicin group compared to the control (DEN+) group. Additional evidence that capsaicin treatment caused apoptosis in HPCs along the bile duct was provided by the tunnel experiment, which demonstrated that apoptosis only surrounded the tubular structure. Furthermore, through the simultaneous labelling of Caspase-3 and EpCAM, researchers were able to show that capsaicin caused HPCs in the DEN-HCC rat model to undergo apoptosis32.
#
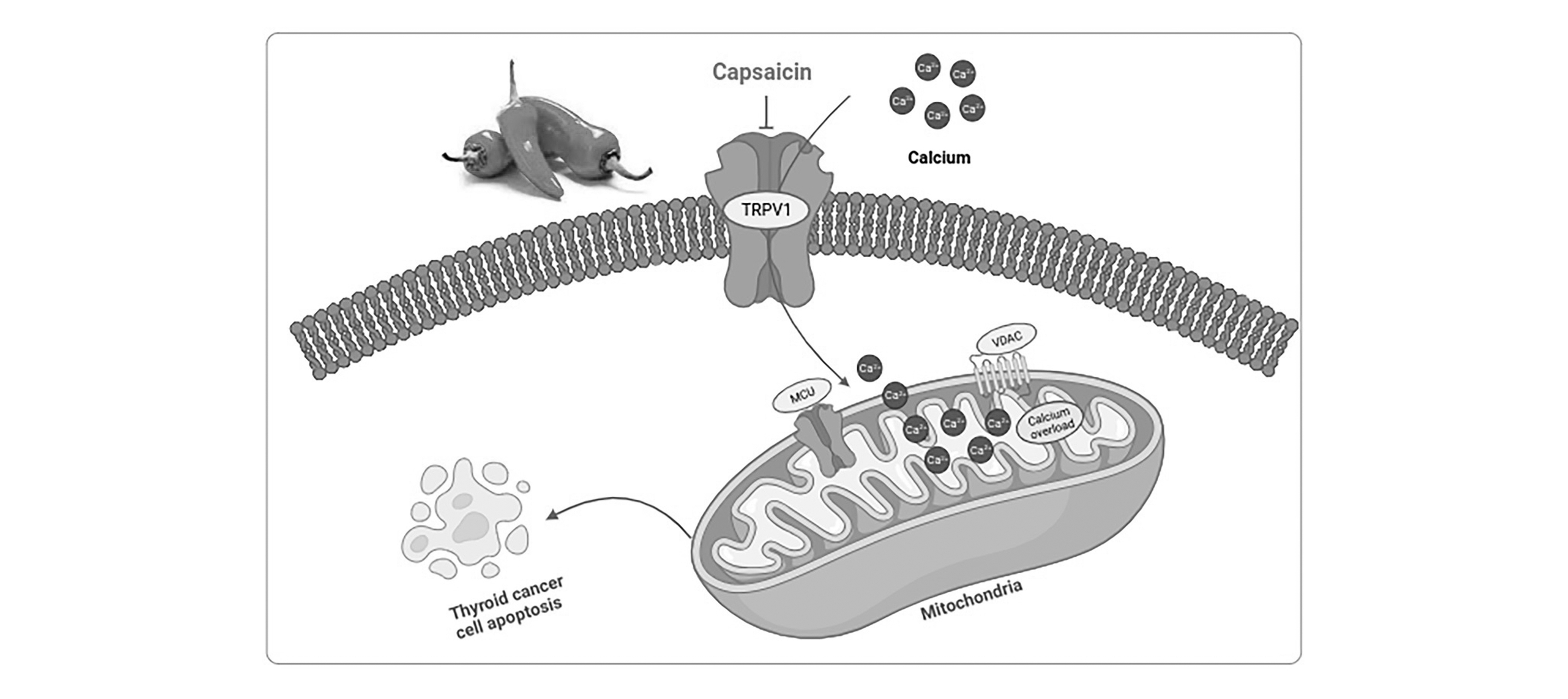
Thyroid Cancer
TRPV1 was shown to be substantially expressed in PTC BCPAP cells, and in ATC cells, capsaicin temporarily increases the cytosolic Ca2+ level. In the current investigation, four additional ATC cell lines and a non-tumorigenic human thyroid follicular cell line were used to further evaluate the expression of the TRPV1 channel by means of reverse transcriptase polymerase chain reaction (RT-PCR) and western blotting. TRPV1 mRNA expression levels increased in ATC 8505 C, FRO, and C643 cells as compared to Nthy-ori-3.1 cells. While remaining similar in 8505 C cells to those in Nthy-ori-3.1 cells in 8505 C cells, TRPV1 protein levels also increased in FRO and C643 cells. In 8305 C and C643 cells, TRPV1's baseline transcriptional expression was nevertheless kept at a low level. The 8505 C and FRO cell lines were selected by the researchers for use in further research analyses [35].
It is commonly known that intracellular calcium may be markedly increased by the strong TRPV1 agonist CAP. Thus, researchers measured the intracellular calcium content using a fluorescence spectrophotometer following the staining of Fluo3-AM. Fluo3 showed a significant and time-dependent increase in fluorescence intensity after being treated with 200 mM CAP. In order to ascertain the potential involvement of TRPV1, the particular receptor for CAP, in the CAP-induced calcium influx are shown in [Fig. 7], the 10 mM capsaicin (CAP), an antagonist of TRPV1, was pretreated in 8505 C cells. As expected, the TRPV1 inhibition dramatically reduced the CAP-induced calcium influx. Furthermore, the calcium chelator BAPTA-AM prevented the rise of calcium in the cytoplasm. Additionally, flow cytometry data demonstrated that BAPTA and CPZ slightly reduced the rapid intracellular calcium buildup that CAP caused in ATC 8505 C and FRO cells. Our findings suggested that the TRPV1 channel in ATC cells may be used by CAP to temporarily increase calcium entry [36].
#
#
Conclusion
In conclusion, the comprehensive review of capsaicin's involvement in the prevention and treatment of cancer uncovers a fascinating and complex scenario. The diverse effects of capsaicin on cancer cells, such as its capacity to trigger programmed cell death, hinder cell division, obstruct the formation of new blood vessels, and perhaps prevent the spread of cancer to other parts of the body, provide a hopeful basis for further investigation. The data confirming capsaicin's ability to induce apoptosis is remarkable since this mechanism offers a focused method for destroying cancer cells. Capsaicin can activate programmed cell death pathways, which makes it a promising candidate for explicitly targeting and eliminating aberrant cells, an essential factor in cancer treatment. The inhibitory effects of capsaicin on cell proliferation further enhance its potential as a therapeutic agent for cancer. Suppressing the unregulated proliferation of cancer cells is a crucial factor in impeding the advancement of tumors, and the influence of capsaicin on the regulation of cell division provides opportunities for exploring its precise molecular processes. The literature review also emphasizes capsaicin's ability to disrupt angiogenesis, a crucial factor in tumor formation. Capsaicin may impede the development of new blood vessels, which may restrict the provision of nutrients to tumors, impeding their growth and survival. Moreover, the reported capacity of capsaicin to hinder the movement and infiltration of cancer cells implies its involvement in limiting the spread of cancer to other parts of the body, which is crucial for enhancing the outlook of the disease. Essentially, the thorough analysis highlights capsaicin's promising prospects in preventing and treating cancer.
#
#
Conflict of Interest
The authors declare that they have no conflict of interest.
Acknowledgment
The authors would like to thank M.S. Ramaiah University of Applied Sciences for supporting this project by providing seed money grant (ORISG/FPH/005/2023). The authors extend their appreciation to the Deanship of Scientific Research at King Khalid University for funding this work through Large Group Research Project under grant number (RGP2/131/44).
-
References
- 1 Kunjiappan S, Sankaranarayanan M, Kumar BK. et al. Capsaicin-loaded solid lipid nanoparticles: Design, biodistribution, in silico modeling and in vitro cytotoxicity evaluation. Nanotechnology 2020; 32: 095101
- 2 Clark R, Lee SH. Anticancer properties of capsaicin against human cancer. Anticancer Research 2016; 36: 837-843
- 3 Kang JH, Kim CS, Han IS. et al. Capsaicin, a spicy component of hot peppers, modulates adipokine gene expression and protein release from obese-mouse adipose tissues and isolated adipocytes, and suppresses the inflammatory responses of adipose tissue macrophages. FEBS Letters 2007; 581: 4389-4396
- 4 Sánchez BG, Bort A, Mora-Rodríguez JM. et al. The Natural Chemotherapeutic Capsaicin Activates AMPK through LKB1 Kinase and TRPV1 Receptors in Prostate Cancer Cells. Pharmaceutics 2022; 14: 329
- 5 Sharma SK, Vij AS, Sharma M. Mechanisms and clinical uses of capsaicin. European Journal of Pharmacology 2013; 720: 55-62
- 6 Toh C, Lee T, Kiang A. The pharmacological actions of capsaicin and analogues. British Journal of Pharmacology and Chemotherapy 1955; 10: 175
- 7 Lang Y, Yanagawa S, Sasanuma T. et al. A gene encoding a putative acyl-transferase involved in pungency of Capsicum. Breeding Science 2006; 56: 55-62
- 8 Reilly CA, Ehlhardt WJ, Jackson DA. et al. Metabolism of capsaicin by cytochrome P450 produces novel dehydrogenated metabolites and decreases cytotoxicity to lung and liver cells. Chemical Research in Toxicology 2003; 16: 336-349
- 9 Raisinghani M, Pabbidi RM, Premkumar LS. Activation of transient receptor potential vanilloid 1 (TRPV1) by resiniferatoxin. The Journal of Physiology 2005; 567: 771-786
- 10 Ziglioli F, Frattini A, Maestroni U. et al. Vanilloid-mediated apoptosis in prostate cancer cells through a TRPV-1 dependent and a TRPV-1-independent mechanism. Acta bio-medica: Atenei Parmensis 2009; 80: 13-20
- 11 Zhang JH, Lai FJ, Chen H. et al. Involvement of the phosphoinositide 3‑kinase/Akt pathway in apoptosis induced by capsaicin in the human pancreatic cancer cell line PANC-1. Oncology Letters 2013; 5: 43-48
- 12 Pramanik KC, Boreddy SR, Srivastava SK. Role of mitochondrial electron transport chain complexes in capsaicin mediated oxidative stress leading to apoptosis in pancreatic cancer cells. PloS one 2011; 6: e20151
- 13 Mori A, Lehmann SR, O'Kelly J. et al. Capsaicin, a component of red peppers, inhibits the growth of androgen-independent, p53 mutant prostate cancer cells. Cancer Research 2006; 66: 3222-3229
- 14 Moon DO, Kang CH, Kang SH. et al. Capsaicin sensitizes TRAIL-induced apoptosis through Sp1-mediated DR5 up-regulation: Involvement of Ca2+ influx. Toxicology and Applied Pharmacology 2012; 259: 87-95
- 15 Chang H, Chen S, Chien S. et al. Capsaicin may induce breast cancer cell death through apoptosis-inducing factor involving mitochondrial dysfunction. Human & Experimental Toxicology 2011; 30: 1657-1665
- 16 de-Sá-Júnior PL, Pasqualoto KFM, Ferreira AK. et al. RPF101, a new capsaicin-like analogue, disrupts the microtubule network accompanied by arrest in the G2/M phase, inducing apoptosis and mitotic catastrophe in the MCF-7 breast cancer cells. Toxicology and Applied Pharmacology 2013; 266: 385-398
- 17 Lu HF, Chen YL, Yang JS. et al. Antitumor activity of capsaicin on human colon cancer cells in vitro and colo 205 tumor xenografts in vivo. Journal of Agricultural and Food Chemistry 2010; 58: 12999-13005
- 18 Wang HM, Chuang SM, Su YC. et al. Down-regulation of tumor-associated NADH oxidase, tNOX (ENOX2), enhances capsaicin-induced inhibition of gastric cancer cell growth. Cell Biochemistry and Biophysics 2011; 61: 355-366
- 19 Tsou MF, Lu H-F, Chen SC. et al. Involvement of Bax, Bcl-2, Ca2+ and caspase-3 in capsaicin-induced apoptosis of human leukemia HL-60 cells. Anticancer Research 2006; 26: 1965-1971
- 20 Dimmeler S, Fleming I, Fisslthaler B. et al. Activation of nitric oxide synthase in endothelial cells by Akt-dependent phosphorylation. Nature 1999; 399: 601-605
- 21 Morbidelli L, Chang C-H, Douglas JG. et al. Nitric oxide mediates mitogenic effect of VEGF on coronary venular endothelium. American Journal of Physiology-Heart and Circulatory Physiology 1996; 270: H411-H415
- 22 Papapetropoulos A, García-Cardeña G, Madri JA. et al. Nitric oxide production contributes to the angiogenic properties of vascular endothelial growth factor in human endothelial cells. The Journal of Clinical Investigation 1997; 100: 3131-3139
- 23 Min JK, Han KY, Kim EC. et al. Capsaicin inhibits in vitro and in vivo angiogenesis. Cancer Research 2004; 64: 644-651
- 24 Baselga J. The EGF receptor family as targets for breast cancer therapy. Breast Cancer Research 2000; 2 S. 13
- 25 Krueger JS, Keshamouni VG, Atanaskova N. et al. Temporal and quantitative regulation of mitogen-activated protein kinase (MAPK) modulates cell motility and invasion. Oncogene 2001; 20: 4209-4218
- 26 Thoennissen N, O'kelly J, Lu D. et al. Capsaicin causes cell-cycle arrest and apoptosis in ER-positive and-negative breast cancer cells by modulating the EGFR/HER-2 pathway. Oncogene 2010; 29: 285-296
- 27 Yoon JH, Ahn SG, Lee BH. et al. Role of autophagy in chemoresistance: regulation of the ATM-mediated DNA-damage signaling pathway through activation of DNA–PKcs and PARP-1. Biochemical Pharmacology 2012; 83: 747-757
- 28 Gobeil S, Boucher C, Nadeau D. et al. Characterization of the necrotic cleavage of poly (ADP-ribose) polymerase (PARP-1): implication of lysosomal proteases. Cell Death & Differentiation 2001; 8: 588-594
- 29 Virag L, Szabó C. The therapeutic potential of poly (ADP-ribose) polymerase inhibitors. Pharmacological Reviews 2002; 54: 375-429
- 30 Di Giovanni S, Mirabella M, Papacci M. et al. Apoptosis and ROS detoxification enzymes correlate with cytochrome c oxidase deficiency in mitochondrial encephalomyopathies. Molecular and Cellular Neuroscience 2001; 17: 696-705
- 31 Lee MG, Lee KT, Chi SG. et al. Constunolide induces apoptosis by ROS-mediated mitochondrial permeability transition and cytochrome c release. Biological and Pharmaceutical Bulletin 2001; 24: 303-306
- 32 Semenza GL. Targeting HIF-1 for cancer therapy. Nature Reviews Cancer 2003; 3: 721-732
- 33 Semenza GL. Hypoxia-inducible factors in physiology and medicine. Cell 2012; 148: 399-408
- 34 Han TH, Park MK, Nakamura H. et al. Capsaicin inhibits HIF-1α accumulation through suppression of mitochondrial respiration in lung cancer cells. Biomedicine & Pharmacotherapy 2022; 146: 112500
- 35 Wang HP, Pu XY, Wang XH. Distribution profiles of transient receptor potential melastatin-related and vanilloid-related channels in prostatic tissue in rat. Asian Journal of Andrology 2007; 9: 634-640
- 36 Xu S, Cheng X, Wu J. et al. Capsaicin restores sodium iodine symporter-mediated radioiodine uptake through bypassing canonical TSH-TSHR pathway in anaplastic thyroid carcinoma cells. Journal of Molecular Cell Biology 2021; 13: 791-807
Correspondence
Publication History
Received: 12 March 2024
Accepted: 16 April 2024
Article published online:
10 May 2024
© 2024. Thieme. All rights reserved.
Georg Thieme Verlag KG
Rüdigerstraße 14, 70469 Stuttgart,
Germany
-
References
- 1 Kunjiappan S, Sankaranarayanan M, Kumar BK. et al. Capsaicin-loaded solid lipid nanoparticles: Design, biodistribution, in silico modeling and in vitro cytotoxicity evaluation. Nanotechnology 2020; 32: 095101
- 2 Clark R, Lee SH. Anticancer properties of capsaicin against human cancer. Anticancer Research 2016; 36: 837-843
- 3 Kang JH, Kim CS, Han IS. et al. Capsaicin, a spicy component of hot peppers, modulates adipokine gene expression and protein release from obese-mouse adipose tissues and isolated adipocytes, and suppresses the inflammatory responses of adipose tissue macrophages. FEBS Letters 2007; 581: 4389-4396
- 4 Sánchez BG, Bort A, Mora-Rodríguez JM. et al. The Natural Chemotherapeutic Capsaicin Activates AMPK through LKB1 Kinase and TRPV1 Receptors in Prostate Cancer Cells. Pharmaceutics 2022; 14: 329
- 5 Sharma SK, Vij AS, Sharma M. Mechanisms and clinical uses of capsaicin. European Journal of Pharmacology 2013; 720: 55-62
- 6 Toh C, Lee T, Kiang A. The pharmacological actions of capsaicin and analogues. British Journal of Pharmacology and Chemotherapy 1955; 10: 175
- 7 Lang Y, Yanagawa S, Sasanuma T. et al. A gene encoding a putative acyl-transferase involved in pungency of Capsicum. Breeding Science 2006; 56: 55-62
- 8 Reilly CA, Ehlhardt WJ, Jackson DA. et al. Metabolism of capsaicin by cytochrome P450 produces novel dehydrogenated metabolites and decreases cytotoxicity to lung and liver cells. Chemical Research in Toxicology 2003; 16: 336-349
- 9 Raisinghani M, Pabbidi RM, Premkumar LS. Activation of transient receptor potential vanilloid 1 (TRPV1) by resiniferatoxin. The Journal of Physiology 2005; 567: 771-786
- 10 Ziglioli F, Frattini A, Maestroni U. et al. Vanilloid-mediated apoptosis in prostate cancer cells through a TRPV-1 dependent and a TRPV-1-independent mechanism. Acta bio-medica: Atenei Parmensis 2009; 80: 13-20
- 11 Zhang JH, Lai FJ, Chen H. et al. Involvement of the phosphoinositide 3‑kinase/Akt pathway in apoptosis induced by capsaicin in the human pancreatic cancer cell line PANC-1. Oncology Letters 2013; 5: 43-48
- 12 Pramanik KC, Boreddy SR, Srivastava SK. Role of mitochondrial electron transport chain complexes in capsaicin mediated oxidative stress leading to apoptosis in pancreatic cancer cells. PloS one 2011; 6: e20151
- 13 Mori A, Lehmann SR, O'Kelly J. et al. Capsaicin, a component of red peppers, inhibits the growth of androgen-independent, p53 mutant prostate cancer cells. Cancer Research 2006; 66: 3222-3229
- 14 Moon DO, Kang CH, Kang SH. et al. Capsaicin sensitizes TRAIL-induced apoptosis through Sp1-mediated DR5 up-regulation: Involvement of Ca2+ influx. Toxicology and Applied Pharmacology 2012; 259: 87-95
- 15 Chang H, Chen S, Chien S. et al. Capsaicin may induce breast cancer cell death through apoptosis-inducing factor involving mitochondrial dysfunction. Human & Experimental Toxicology 2011; 30: 1657-1665
- 16 de-Sá-Júnior PL, Pasqualoto KFM, Ferreira AK. et al. RPF101, a new capsaicin-like analogue, disrupts the microtubule network accompanied by arrest in the G2/M phase, inducing apoptosis and mitotic catastrophe in the MCF-7 breast cancer cells. Toxicology and Applied Pharmacology 2013; 266: 385-398
- 17 Lu HF, Chen YL, Yang JS. et al. Antitumor activity of capsaicin on human colon cancer cells in vitro and colo 205 tumor xenografts in vivo. Journal of Agricultural and Food Chemistry 2010; 58: 12999-13005
- 18 Wang HM, Chuang SM, Su YC. et al. Down-regulation of tumor-associated NADH oxidase, tNOX (ENOX2), enhances capsaicin-induced inhibition of gastric cancer cell growth. Cell Biochemistry and Biophysics 2011; 61: 355-366
- 19 Tsou MF, Lu H-F, Chen SC. et al. Involvement of Bax, Bcl-2, Ca2+ and caspase-3 in capsaicin-induced apoptosis of human leukemia HL-60 cells. Anticancer Research 2006; 26: 1965-1971
- 20 Dimmeler S, Fleming I, Fisslthaler B. et al. Activation of nitric oxide synthase in endothelial cells by Akt-dependent phosphorylation. Nature 1999; 399: 601-605
- 21 Morbidelli L, Chang C-H, Douglas JG. et al. Nitric oxide mediates mitogenic effect of VEGF on coronary venular endothelium. American Journal of Physiology-Heart and Circulatory Physiology 1996; 270: H411-H415
- 22 Papapetropoulos A, García-Cardeña G, Madri JA. et al. Nitric oxide production contributes to the angiogenic properties of vascular endothelial growth factor in human endothelial cells. The Journal of Clinical Investigation 1997; 100: 3131-3139
- 23 Min JK, Han KY, Kim EC. et al. Capsaicin inhibits in vitro and in vivo angiogenesis. Cancer Research 2004; 64: 644-651
- 24 Baselga J. The EGF receptor family as targets for breast cancer therapy. Breast Cancer Research 2000; 2 S. 13
- 25 Krueger JS, Keshamouni VG, Atanaskova N. et al. Temporal and quantitative regulation of mitogen-activated protein kinase (MAPK) modulates cell motility and invasion. Oncogene 2001; 20: 4209-4218
- 26 Thoennissen N, O'kelly J, Lu D. et al. Capsaicin causes cell-cycle arrest and apoptosis in ER-positive and-negative breast cancer cells by modulating the EGFR/HER-2 pathway. Oncogene 2010; 29: 285-296
- 27 Yoon JH, Ahn SG, Lee BH. et al. Role of autophagy in chemoresistance: regulation of the ATM-mediated DNA-damage signaling pathway through activation of DNA–PKcs and PARP-1. Biochemical Pharmacology 2012; 83: 747-757
- 28 Gobeil S, Boucher C, Nadeau D. et al. Characterization of the necrotic cleavage of poly (ADP-ribose) polymerase (PARP-1): implication of lysosomal proteases. Cell Death & Differentiation 2001; 8: 588-594
- 29 Virag L, Szabó C. The therapeutic potential of poly (ADP-ribose) polymerase inhibitors. Pharmacological Reviews 2002; 54: 375-429
- 30 Di Giovanni S, Mirabella M, Papacci M. et al. Apoptosis and ROS detoxification enzymes correlate with cytochrome c oxidase deficiency in mitochondrial encephalomyopathies. Molecular and Cellular Neuroscience 2001; 17: 696-705
- 31 Lee MG, Lee KT, Chi SG. et al. Constunolide induces apoptosis by ROS-mediated mitochondrial permeability transition and cytochrome c release. Biological and Pharmaceutical Bulletin 2001; 24: 303-306
- 32 Semenza GL. Targeting HIF-1 for cancer therapy. Nature Reviews Cancer 2003; 3: 721-732
- 33 Semenza GL. Hypoxia-inducible factors in physiology and medicine. Cell 2012; 148: 399-408
- 34 Han TH, Park MK, Nakamura H. et al. Capsaicin inhibits HIF-1α accumulation through suppression of mitochondrial respiration in lung cancer cells. Biomedicine & Pharmacotherapy 2022; 146: 112500
- 35 Wang HP, Pu XY, Wang XH. Distribution profiles of transient receptor potential melastatin-related and vanilloid-related channels in prostatic tissue in rat. Asian Journal of Andrology 2007; 9: 634-640
- 36 Xu S, Cheng X, Wu J. et al. Capsaicin restores sodium iodine symporter-mediated radioiodine uptake through bypassing canonical TSH-TSHR pathway in anaplastic thyroid carcinoma cells. Journal of Molecular Cell Biology 2021; 13: 791-807