
RSS-Feed abonnieren
DOI: 10.1055/a-1320-4610
The Potential of Biologically Active Brazilian Plant Species as a Strategy to Search for Molecular Models for Mosquito Control
Gefördert durch: Fundação de Empreendimentos Científicos e Tecnológicos 120/2017
Gefördert durch: Fundação de Amparo à Pesquisa do Estado de São Paulo 2013/07600-3
Gefördert durch: Fundação de Amparo à Pesquisa do Estado de São Paulo 2014/50926-0
Gefördert durch: Fundação de Amparo à Pesquisa do Estado de São Paulo 2019/05967-3

- Abstract
- Introduction
- Natural Products from Brazilian Biodiversity as a Source of New Models for Aedes aegypti Control
- Molecular Targets for the Design of Compounds to Control Aedes aegypti
- Conclusions
- Contributorsʼ Statement
- References
Abstract
Natural products are a valuable source of biologically active compounds and continue to play an important role in modern drug discovery due to their great structural diversity and unique biological properties. Brazilian biodiversity is one of the most extensive in the world and could be an effective source of new chemical entities for drug discovery. Mosquitoes are vectors for the transmission of dengue, Zika, chikungunya, yellow fever, and many other diseases of public health importance. These diseases have a major impact on tropical and subtropical countries, and their incidence has increased dramatically in recent decades, reaching billions of people at risk worldwide. The prevention of these diseases is mainly through vector control, which is becoming more difficult because of the emergence of resistant mosquito populations to the chemical insecticides. Strategies to provide efficient and safe vector control are needed, and secondary metabolites from plant species from the Brazilian biodiversity, especially Cerrado, that are biologically active for mosquito control are herein highlighted. Also, this is a literature revision of targets as insights to promote advances in the task of developing active compounds for vector control. In view of the expansion and occurrence of arboviruses diseases worldwide, scientific reviews on bioactive natural products are important to provide molecular models for vector control and contribute with effective measures to reduce their incidence.
#
Key words
Cerrado - Brazilian hotspot - natural products - molecular targets - arbovirus - Aedes aegyptiAbbreviations
Introduction
Natural products still stand out among the 1394 small-molecule approved drugs introduced in the market as pharmaceuticals during the period 1981 – 2019 in which more than 65% are derived from natural products or with an inspired structure [1]. Tropical ecosystems are particularly rich in chemistry diversity [2] because of the large number of different species present and the interactions among these species. Brazil has a huge biodiversity accounting for 20% of all living species [3]. Therefore, the search for natural products in plants from Brazilian biomes, a rich source of biologically active compounds [4], is an important source of new chemical entities in drug discovery. The Cerrado is a genuinely Brazilian savanna that covers about 2 million km2 or 21% of the Brazilian territory [4] and is considered to be a hotspot of biodiversity with a high concentration of species, e.g., more than 10 000 species of plants, which means high levels of biodiversity [5]. However, due to the expansion of agricultural activities as well as extensive deforestation, several species of the Cerrado are at risk of extinction [3], [6]. This is the second largest Brazilian biome, with singular characteristics of plant morphologies, soil, climate, and landscape [7]. Its flora accounts for 40% of endemic species, being the most biodiverse tropical savanna in the world [7]. Together with the Atlantic Forest, this is the most commonly reported biome in the NuBBEDB [8]. The accelerate loss of Cerrado biodiversity will disturb the biome, and its ecological imbalance will increase the risk of infectious diseases caused by mosquitoes. Natural products from Brazilian biomes, especially from Cerrado, could be interesting models for insecticide discovery, which will be addressed in this review.
Mosquitoes are important arbovirus vectors distributed in more than 100 genera from the order Diptera, family Culicidae. Aedes, Culex, and Anopheles genera are the main vectors of several arboviruses that cause diseases of great public health impact [9]. The genus Anopheles is responsible for transmiting the protozoa Plasmodium falciparum that causes malaria. This genus includes more than 400 species and approximately 70 of these are reported to transmit malaria parasites. Most common vectors of malaria by regions are An. gambiae in Africa, An. Farauti in Asia, An. atroparvus in Europe/Middle East, and An. darlingi in Brazil [10]. Mosquitoes from the Culex genus are vectors of arboviruses such as the agents that cause West Nile fever and Japanese encephalitis, transmitting diseases to humans, birds, and other animals. Culex species are spread worldwide. Culex pipiens and Culex quinquefasciatus are prevalent in urbanized areas, while Culex tarsalis is mostly found in rural areas. [9], [11]. The mosquito Aedes aegypti is the main vector for the arboviruses that cause dengue, Zika, chikungunya [12], and yellow fever [9], diseases that are of great impact on worldwide health.
Vector control is currently one of the main strategies to prevent these diseases. This review deals with plant species explored as a sustainable source for vector control of Ae. aegypti. This is also an extensive literature review of targets as insights to promote advances in the task of developing products for mosquito control. Such products require optimization of pharmacodynamic and pharmacokinetic properties that might be tackled by a medicinal chemistry approach and is herein discussed. Although the mosquito species main focus of this review is Ae. aegypti, the bioactive agents could be further explored in research for developing insecticides for many mosquito species.
Ae. aegypti is now distributed in almost all continents, putting billions of people at risk of the diseases transmitted by this vector [13]. Additionally, these diseases have a secondary vector and can also be transmited by Aedes Albopictus, which is present in Asia and parts of the Americas [13]. To contextualize the impact of the diseases transmited by these mosquitoes, dengue is caused by four serotypes of dengue virus (DENV1 – 4), has been identified in 128 countries, and its geographical distribution has increased in the past decades [14]. The global incidence of dengue has increased dramatically in recent decades and about half of the worldʼs population, (est. 3.9 billion people) is at risk [14], [15]. An estimate indicates 390 million dengue infections every year worldwide [15], [16]. In Brazil, the incidence of dengue has grown considerably; in 2018, 265 934 cases (155 deaths) were reported, in 2019, there were 2 226 914 (789 deaths) cases, and in only 16 weeks of 2020, there were 794 565 cases of dengue and 181 deaths [17]. ZIKV transmission is primarily via the mosquito Ae. aegypti, but it is also transmitted during pregnancy, sexual contact, transfusion of blood, and organ transplantation [18]. The first recorded incidence of ZIKV disease was reported on the Island of Yap in 2007, followed by cases in several other countries. The ZIKV infection has been associated with central nervous system diseases, congenital microcephaly in children of women who had Zika, and pregnancy complications such as fetal loss, stillbirth, and preterm birth [18], [19]. CHIKV infection is an economical problem for many countries and has been identified in Asia, Africa, Europe, and the Americas, with over 2 million cases being reported since 2005 [20].
There is still no specific drug available to treat dengue, as well as neither vaccines nor drugs to prevent or treat Zika or chikungunya [13], [21]. The first dengue vaccine, Dengvaxia (CYD-TDV) developed by Sanofi Pasteur, is an attenuated tetravalent product licensed in 2015, and approved in 20 countries for use in people aged 9 – 45 years living in endemic areas [22], [23], [24]. However, its use is limited because of an increased risk of more severe forms of dengue due to the vaccine [24]. The prophylaxis of these diseases is mainly through vector control [15]. Mosquito control is a challenge as resistance has developed in mosquitoes and there is a constant use of insecticides with high toxicity, which has become a major environmental concern. In addition, there is a high density of human populations and a general lack of resources in most endemic countries [25].
The most widespread method for controlling disease vectors is the application of chemical insecticides. Many of these have been successful against Ae. aegypti larvae and adults such as organophosphates, organochlorines, and pyrethroids. However, current chemical agents are not selective and can be toxic to non-target organisms. In addition, considering the insecticide resistance in several areas of the world, its use has been facing problems and the search for new alternatives for the control of disease vectors is needed [26]. Resistance creates yet another difficulty in vector control and raises questions about environmental safety and toxicity to other species, including humans. In Brazil, for example, the control of Ae. aegypti began in 1947 with the use of the organochloride DDT. In 1955, the mosquito was eradicated, but was reintroduced in Brazil in 1967, and temephos was then used for its control because it was already resistant to DDT. In 1973, the mosquito was eradicated for the second time, but reintroduced in 1976 and not eradicated since [27]. This showed that insecticides can successfully control mosquito vectors to the point of complete eradication. However, new products are constantly needed to cope with reintroductions and insecticide resistance.
#
Natural Products from Brazilian Biodiversity as a Source of New Models for Aedes aegypti Control
Natural products are a valuable source of chemical compounds that can be used for the development of new insecticides. Plant-based insecticides have been widely used in agricultural pest control and against vectors of insects that impact public health. Plant extracts are expected to be biodegradable and pose a low risk to other living organisms. However, toxicity tests should always be conducted to ensure the safety of these natural products [26]. Considering the potential of the Brazilian biodiversity in the discovery of new environmentally safe biological secondary metabolites for mosquito control, studies on Brazilian plants are herein presented to show their potential for the development of new mosquito control innovation. [Table 1] lists the larvicidal activity (LC50) of species from 32 Brazilian plant families against Ae. aegypti. The search was made in the SciFinder database (up to 2020) using the term “Aedes” and refined by the words “Brazil” and “plant” and the PubMed database (up to 2020) using the terms “Aedes”, “Brazil” “plant”, and “LC50”. All articles in Portuguese and English were selected and the criterion of inclusion was the report of the LC50 values (24 – 48 h) of native Brazilian plant extracts against Ae. aegypti. When several life cycle forms were active, we report the larvicidal activity, which usually shows the lower LC50.
|
Family |
Species |
Plant part |
Extract, oil, e. o.1 |
LC50 (24 h) ppm |
Ref. |
|---|---|---|---|---|---|
|
1e. o.: essential oil, 248 h, 3chloroform partition, 4hexane fraction, 5nanoemulsion, 6pupicidal activity, 7ethyl acetate fraction, 8fraction from the ethanolic extract, n. a.: not available |
|||||
|
Anacardiaceae |
Anacardium humile A. St.-Hil. |
leaves |
e. o. |
21 |
[28] |
|
Anacardium occidentalis L. |
n. a. |
oil |
142 |
||
|
Myracrodruon urundeuva Fr. Allemao |
seeds |
ethanol |
15 |
[31] |
|
|
leaves |
water |
202 |
[32] |
||
|
Schinopsis brasiliensis Engler |
seeds |
ethanol |
661 |
||
|
stem bark |
ethanol 90% |
3452, 3 |
|||
|
Schinus terebenthifolia Raddi |
fruits |
e. o. |
172 – 344 |
||
|
Annonaceae |
Annona crassiflora Mart |
root bark |
ethanol |
0.712 |
[35] |
|
root wood |
8.92 |
||||
|
stem |
162 |
||||
|
Annona glabra L. |
seeds |
ethanol |
0.062 |
[35] |
|
|
stem |
27 |
[29] |
|||
|
Annona muricata L. |
roots |
ethanol |
422 |
[35] |
|
|
seeds |
ethanol |
94 |
|||
|
Annona squamosa L. |
roots |
ethanol |
322 |
[35] |
|
|
leaves |
1692 |
||||
|
seeds |
52 |
||||
|
Asteraceae |
Acmella oleracea L. |
leaves |
ethanol |
1454 |
[38] |
|
Ageratum conyzoides L. |
n. a. |
e. o. |
1482 |
||
|
Artemisia vulgaris L. |
leaves |
e. o. |
114 |
||
|
Baccharis reticularia DC. |
leaves |
e. o. |
1182, 5 |
[40] |
|
|
Tagetes erecta L. |
stem |
e. o. |
80 |
[41] |
|
|
Bignoniaceae |
Tabebuia avellanedae Lorentz ex Griseb. |
trunk bark |
ethanol |
442 |
[42] |
|
wood |
acetone |
1002 |
[43] |
||
|
ethyl acetate |
1512 |
||||
|
Boraginaceae |
Auxemma glazioviana Taub. |
heartwood |
e. o. |
2 – 3 |
[44] |
|
Cordia curassavica (Jacq.) Roem. & Schult. |
leaves |
e. o. |
97 |
[45] |
|
|
Cordia leucomalloides Taroda |
63 |
||||
|
Cordia globosa (Jacq.) HBK. |
leaves |
e. o. |
28 |
[46] |
|
|
Burseraceae |
Commiphora leptophloeos (Mart.) J. B. Gillett |
leaves |
e. o. |
99 |
[47] |
|
Combretaceae |
Terminalia fagifolia Mart. |
trunk |
ethanol |
373 |
[42] |
|
Connaraceae |
Connarus detersus Planch. |
seeds |
ethanol |
2166 |
|
|
Rourea doniana Baker |
leaves |
chloroform |
171 |
[48] |
|
|
stem |
hexane |
12 |
|||
|
Convolvulaceae |
Merremia aegyptia (L.) Urb. |
leaves |
acetone |
120 |
[48] |
|
hexane |
144 |
||||
|
Euphorbiaceae |
Croton argyrophylloides Müll. Arg. |
leaves |
e. o. |
95 |
|
|
Croton heliotropiifolius Kunth |
leaves |
e. o. |
544 |
[51] |
|
|
Croton pulegiodorus Baill. |
159 |
||||
|
Croton nepetifolius Baill. |
leaves |
e. o. |
66 |
[49] |
|
|
Croton rhamnifolioides Pax & K. Hoffm. |
leaves |
e. o. |
122 |
[52] |
|
|
Croton regelianus Müll. Arg. |
leaves |
e. o. |
24 |
[53] |
|
|
Croton sonderianus Müll. Arg. |
leaves |
e. o. |
55 |
[49] |
|
|
Croton tetradenius Baill. |
leaves |
e. o. |
152 |
[54] |
|
|
Croton zehntneri Pax & K. Hoffm. |
leaves |
e. o. |
26 |
[49] |
|
|
Fabaceae |
Albizia polyantha (A. Spreng.) G. P. Lewis |
stem |
ethanol |
547 |
[42] |
|
Anadenanthera macrocarpa (Benth.) Brenan |
seeds |
water |
430 |
[55] |
|
|
Copaifera langsdorffii Desf. |
n. a. |
oil |
412 |
[29] |
|
|
Copaifera multijuga Hayne |
bark |
ethanol |
81 |
[56] |
|
|
leaves |
ethanol |
166 |
|||
|
oil-resin |
o. e. |
18 |
|||
|
Dalbergia brasiliensis Vogel |
leaves |
ethanol |
247 |
||
|
Derris urucu (Killip & A. C. Sm.) J. F. Macbr. |
root |
ethanol |
18 |
[58] |
|
|
Dioclea megacarpa Rolfe |
seeds |
ethanol |
206 |
||
|
Enterolobium contortisiliquum (Vell.) Morong. |
seeds |
ethanol |
3696 |
||
|
Hymenaea courbaril L. |
fruit peel |
e. o. |
15 |
[59] |
|
|
Luetzelburgia auriculate (Allemão) Ducke |
seeds |
ethanol |
8.66 |
||
|
Ormosia arborea (Vell.) Harms |
leaves |
ethanol |
238 |
||
|
seeds |
ethanol |
111 |
|||
|
Parkia platycephala Benth. |
seeds |
ethanol |
136 |
[31] |
|
|
Senna obtusifolia (L.) H. S. Irwin & Barneby |
seeds |
ethanol |
196 |
[31] |
|
|
Tephrosia egregia Sandwith |
stem |
hexane |
13 |
[61] |
|
|
Lamiaceae |
Hyptis martiusii Benth. |
leaves |
e. o. |
18 |
[62] |
|
Hyptis pectinata (L.) Poit. |
leaves |
e. o. |
366 |
[63] |
|
|
Ocimum carnosum (Spreng.) Link & Otto ex Benth. |
inflorescences |
e. o. |
109 |
[64] |
|
|
Ocimum campechianum Mill. |
leaves |
e. o. |
81 |
[64] |
|
|
Lauraceae |
Licaria puchury-major (Mart.) Kosterm. |
seeds |
e. o. |
99 |
[65] |
|
Ocotea velloziana (Meisn.) Mez |
trunk bark |
ethanol |
213 |
[42] |
|
|
Leguminosae |
Copaifera reticulata Ducke |
trunk |
oil-resin |
8.9 |
[66] |
|
Bauhinia acuruana (Moric) |
leaves |
e. o. |
56 |
[67] |
|
|
Derris sp. |
roots |
ethanol |
4.82 |
[35] |
|
|
Erythrina mulungu Mart. ex Benth. |
stem bark |
ethanol |
372 |
[35] |
|
|
Pterodon polygalaeflorus Benth |
seeds |
ethanol |
202 |
[35] |
|
|
Malvaceae |
Helicteres velutina K. Schum. |
roots |
ethanol |
171 |
[68] |
|
stem |
ethanol |
139 |
|||
|
Waltheria viscosissima A. St.-Hil |
roots |
ethanol |
5 |
[69] |
|
|
Meliaceae |
Carapa guianensis Aubl. |
n. a. |
oil |
16 – 572 |
|
|
Guarea kunthiana A. Juss. |
seeds |
ethanol |
169 |
[71] |
|
|
Guarea humaitensis T. D. Penn. |
leaves and branches |
e. o. |
49 |
[72] |
|
|
Guarea scabra A. Juss. |
leaves and branches |
e. o. |
99 |
[72] |
|
|
Guarea sylvatica C. DC. |
leaves and branches |
e. o. |
118 |
[72] |
|
|
Myrtaceae |
Eugenia brejoensis Mazine |
leaves |
e. o. |
214 |
[73] |
|
Eugenia candolleana DC. |
leaves |
e. o. |
300 |
[74] |
|
|
Eugenia piauhiensis Vellaff. |
leaves |
e. o. |
230 |
[75] |
|
|
Myrcia sylvatica (G. Mey.) DC. |
leaves |
e. o. |
792 |
[76] |
|
|
Psidium guajava L. |
leaves |
e. o. |
39 – 64 |
[77] |
|
|
Psidium myrsinites Mart. ex DC. |
leaves |
e. o. |
292 |
[75] |
|
|
Passifloraceae |
Turnera ulmifolia L. |
leaves |
ethanol |
242 |
[60] |
|
Pinaceae |
Pinus caribaea Morelet |
leaves |
acetone |
92 |
[78] |
|
Piperaceae |
Piper aduncum L. |
leaves |
e. o. |
290 |
|
|
hexane |
342 |
||||
|
chloroform |
192 |
||||
|
Piper corcovadensis (Miq.) C. DC. |
leaves |
e. o. |
31 |
[80] |
|
|
Piper hispidum Sw. |
leaves |
ethanol |
169 |
[60] |
|
|
chloroform |
567 |
||||
|
Piper klotzschianum (Kunth) C. DC. |
Seeds |
e. o. |
13 |
[81] |
|
|
roots |
10 |
||||
|
Piper marginatum Jacq. |
leaves |
e. o. |
196 |
[82] |
|
|
stem |
146 |
||||
|
inflorescence |
146 |
||||
|
Piper tuberculatum Jacq. |
leaves |
e. o. |
106 |
[39] |
|
|
Ottonia anisum Spreng. |
seeds |
e. o. |
13 |
[83] |
|
|
roots |
e. o. |
10 |
|||
|
Plantaginacea |
Scoparia dulcis L. |
leaves |
ethanol |
83 |
[68] |
|
Poaceae |
Cymbopogon winterianus Jowitt ex Bor |
n. a. |
e. o. |
982 |
[29] |
|
Cymbopogon flexuosus (Nees ex Steud.) Wats. |
leaves |
e. o. |
121 |
[39] |
|
|
Polygonaceae |
Coccoloba mollis Casar. |
stem |
hexane |
137 |
[48] |
|
stem barks |
hexane |
128 |
|||
|
Triplaris americana L. |
stem |
hexane |
117 |
[48] |
|
|
roots |
hexane |
97 |
|||
|
Rhamnaceae |
Ziziphus joazeiro Mart. |
seeds |
ethanol |
1896 |
[31] |
|
Rubiaceae |
Guettarda grazielae M. R. V. Barbosa |
stem |
ethyl acetate |
51 |
[48] |
|
stem bark |
chloroform |
152 |
|||
|
leaves |
hexane8 |
131 |
|||
|
Spermacoce latifolia Aubl. |
leaves |
hexane |
415 |
[60] |
|
|
methanol |
625 |
||||
|
Spermacoce verticillate L. |
stem |
hexane |
115 |
[48] |
|
|
aerial parts |
hexane |
84 |
|||
|
Rutaceae |
Citrus limonia (L.) Osbeck |
fruit peel |
e. o. |
519 |
[84] |
|
Citrus sinensis (L.) Osbeck |
fruit peel |
e. o. |
538 |
[84] |
|
|
Zanthoxylum sp. |
stem |
ethanol |
537 |
[42] |
|
|
leaves |
435 |
||||
|
Sapindaceae |
Talisia esculenta (A. St.-Hil.) Radlk. |
seeds |
ethanol |
4126 |
[31] |
|
Scrophulariaceae |
Stemodia maritima L. |
leaves |
e. o. |
55 |
[85] |
|
stem |
23 |
||||
|
Siparunaceae |
Siparuna guianensis Aubl. |
leaves |
e. o. |
87 |
[86] |
|
Siparuna camporum (Tul.) A. DC. |
leaves |
e. o. |
251 |
[75] |
|
|
Smilacaceae |
Smilax brasiliensis Spreng. |
leaves |
methanol |
986 |
[87] |
|
Solanaceae |
Solanum variabile Mart. |
leaves |
methanol |
188 |
[60] |
|
Verbenaceae |
Lippia alba (Mill.) N. E. Br. ex Britton & P. Wilson |
leaves |
e. o. |
37 |
|
|
Lippia gracilis Schauer |
56 |
||||
|
26 |
|||||
|
Lippia microphylla Cham. |
76 |
||||
|
Lippia nodiflora Cham. |
107 |
||||
|
Lippia sidoides Cham. |
leaves |
e. o. |
19 |
||
|
Lippia pedunculosa Hayek |
leaves |
e. o. |
58 |
[89] |
|
|
Vitex cymose Bertero ex Spreng. |
stem |
ethanol |
875 |
[42] |
|
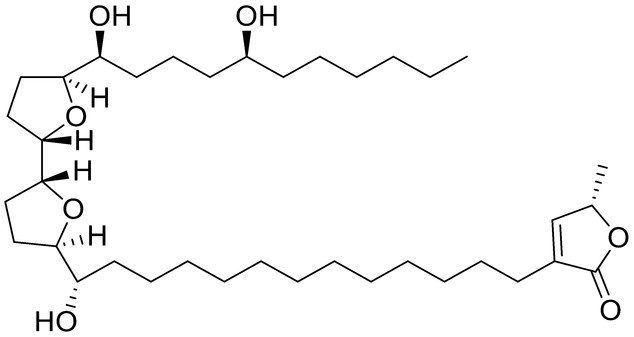
Species of the Annonaceae family have insecticidal properties due to the presence of secondary metabolites compounds known as “annonaceous acetogenins”. These compounds constitute C-35/C-37 natural products derived from fatty acids and show potent and varied biological activities, such as cytotoxic, antitumoral, antimalarial, antifeedant, antimicrobial, pesticidal, and immunosuppressive [45]. Five Annonaceae species collected in the Cerrado region of Mato Grosso-Brazil were extracted with methanol defatted with hexane, methanol defatted with dichloromethane, methanol, hexane, and dichloromethane and tested to evaluate the effect against Ae. aegypti larvae. The species Annona crassiflora, Annona mucosa and Annona coriacea presented high larvicidal activity. The methanol defatted with hexane and hexane extracts from A. coriacea showed 100% activity at 100 µg/mL (LC50 = 7 µg/mL), and the dichloromethane extract had 58.75% activity. Methanol (100 µg/mL) and methanol defatted with hexane (1 mg/mL) from A. mucosa showed 100% activity (LC50 = 10 µg/mL), while the hexane and dichloromethane extracts provided activity higher than 90%. The hexane extract from A. crassiflora (1 mg/mL) provided 91.25% larvae mortality. The phytochemical compounds of Annonaceae can differ between species and solvents used for extraction. These species had high activity larvicide against Ae. aegypti larvae, thus being a potential source to produce new insecticide molecules [90]. Another study tested the toxicity of the acetogenin squamocin (1) ([Table 2]) isolated from A. mucosa seeds collected in Mato Grosso against Ae. aegypti. This compound exhibited good activity with an LC50 = 0.01 µg/mL and LC90 = 0.11 µg/mL [91].
|
Species |
Compound structure |
Stage of mosquito targeted |
Ref. |
|---|---|---|---|
|
Annona mucosa Jacq. |
 squamocin, 1, acetogenin |
Ae. aegypti larvae |
[91] |
|
Duguetia furfuracea (A. St.-Hil.) Saff.; Xylopia aromatica (Lam.) Mart. |
 α-asarole, 2, diterpenoid |
Third-stage Ae. aegypti larvae |
[92] |
 2,4,5-trimethoxy-styrene, 3, diterpenoid |
Third-stage Ae. aegypti larvae |
[92] |
|
 4,2,4,5-trimethoxybenzaldehyde, 4, diterpenoid |
Third-stage Ae. aegypti larvae |
[92] |
|
 ent-labda-8(17),13(16),14-trien-18-oic acid, 5, diterpenoid |
Third-stage Ae. aegypti larvae |
[92] |
|
|
Ocotea velloziana (Meisn.) Mez |
 (+)-dicentrine, 6, alkaloid |
Ae. aegypti larvae |
[42] |
|
Piper aduncum L. |
 1,8-cineole, 7, monoterpenoid |
Ae. aegypti larvae |
[79] |
 α-pinene, 8, monoterpenoid |
Ae. aegypti larvae |
[79] |
|
 β-pinene, 9, monoterpenoid |
Ae. aegypti larvae |
[79] |
|
 trans-ocimene, 10, monoterpenoid |
Ae. aegypti larvae |
[79] |
|
|
Guarea kunthiana A. Juss. |
 melianodiol, 11, protolimonoid |
Ae. aegypti larvae |
[71] |
 meliantriol, 12, protolimonoid |
Ae. aegypti larvae |
[71] |
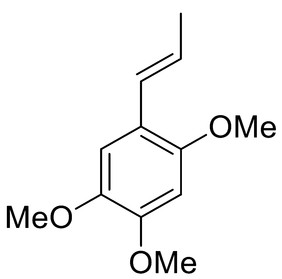
Recent research using a bioguided approach with modern analytical techniques and dereplication strategies showed it was effective in obtaining natural active compounds. Dereplication strategies can identify known compounds in mixtures according to similar fragmentation profiles and masses using datasets. The detection of compounds and the evaluation of the metabolomic profile is possible prior to isolation. The ArboControl Brasil, a program of the Brazilian Ministry of Health, studied the Brazilian Cerrado Plant Extract Bank, revealing active extracts from Annonaceae taxon. Different organs of Cardiopetalum calophyllum, Duguetia furfuracea, Xylopia aromatica, and Xylopia emarginata were prepared with 34 hexane and ethanol crude extracts at a 250 µg/mL concentration in order to test against Ae. aegypti larvae. Only four were active: leaf hexane and root wood ethanol extracts of X. aromatica and root bark and root wood hexane extracts of D. furfuracea. Dereplication strategies were performed on ethyl acetate and methanol extracts and revealed the presence of flavonoids, diterpenes, pheophorbides, and alkaloids. Three phenylpropanoids and one diterpenoid were isolated from D. furfuracea and X. aromatica: α-asarone (2) ([Table 2]), 2,4,5-trimethoxy-styrene (3) ([Table 2]), 2,4,5-trimethoxybenzaldehyde (4) ([Table 2]), and ent-labda-8(17),13(16),14-trien-18-oic acid (5) ([Table 2]). Compound 2 showed 100% mortality of Ae. aegypti larvae at 250 µg/mL. A mixture of 2 and 3 at a ratio of 2 : 1 achieved the highest activity. Extracts from the X. aromatica leaf hexane extract and compound 5 showed that LC50 values decreased (activity increased) with exposure time. These active compounds are promising for the control of Ae. aegypti, showing the potential of the Annonaceae family [92].
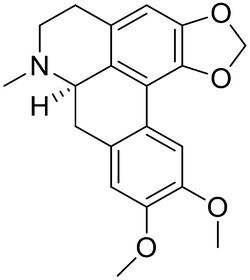
Larvicidal activity against Ae. aegypti was tested on a total of 42 ethanolic extracts from 30 native plants belonging to 18 different families from Pantanal and Cerrado in the region of Mato Grosso do Sul – Brazil. The larvicidal activity was found in six of these plants: Tabebuia avellanedae, Terminalia fagifolia, Ocotea velloziana, Albizia polyantha, Zanthoxylum sp., and Vitex cymosa. Among these species, the most promising result was obtained for the ethanol extract of the trunk bark of O. velloziana (LC50 = 213.70 µg/mL). In the phytochemical investigation, the aporphine alkaloid (+)-dicentrine (6) ([Table 2]) was isolated and characterized as the compound responsible for the larvicidal activity. Other alkaloids have been identified in this species but showed no larvicidal effects. There was no previous information about the activity of the plant or dicentrine against Ae. aegypti and this was the first report in the investigation of this active constituent using a bioassay-guided investigation. The occurrence of dicentrine in several species from neotropical areas of the world can be explored as a potential alternative strategy for the control of Ae. aegypti [42].
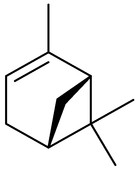
Piper is the largest genus of the family Piperaceae, which is known to be rich in essential oils with insecticidal activity and as insect repellent. Piper aduncum is widely distributed in tropical regions and its essential oil is used in folk medicine to treat respiratory and inflammatory diseases. Studies reported that the essential oil of P. aduncum is active as an insecticide and larvicide, but this activity depends on geographic location from where the plant was collected. This could be due the occurrence of chemotypes and differences in the chemical composition. The larvicide and insecticide action of P. aduncum collected in the Amazon Forest against the larvae and the adult insects of Anopheles marajoana and Ae. aegypti resulted in an LC50 of around 50 ppm. The leaves of this species collected in the Brazilian Cerrado were evaluated and activity was aprox. 280 ppm. The main difference is that the major constituent of the specimen collected in the Amazon Forest is a dillapiole and, in this case, they considered the dillapiole responsible for the biological activity. For the Cerrado specimen, monoterpenes and sesquiterpenes were identified in the essential oil of P. aduncum, and the main monoterpenes, 1,8-cineole (7) ([Table 2]), α-pinene (8) ([Table 2]), β-pinene (9) ([Table 2]), and trans-ocimene (10) ([Table 2]), were identified in great amounts, representing 80% of the essential oil. However, there are differences in the volatile composition of leaves from P. aduncum from different locations that are reported in the literature. Dilapiole was not identified in the Cerrado specimen, hence, the activity observed is due to other metabolites. The evaluation of 1,8-cineole against Ae. aegypti did not provid a larvicidal effect, but the combination of the other monoterpenes increased activity. The collected data suggest that the essential oil showed great larvicidal activity against Ae. aegypti and it can be a new possibility for combating the mosquito due to the components present in the essential oil [79].
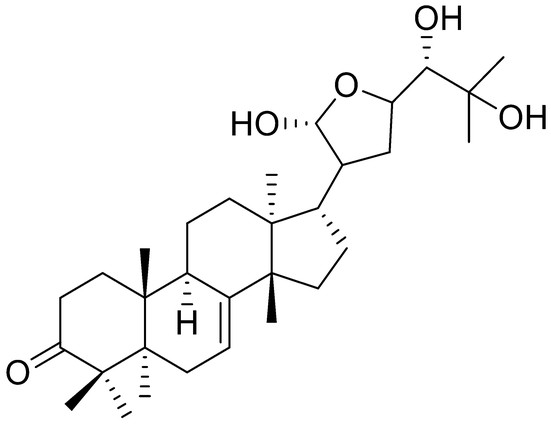
Meliaceae is a family of the genus Guarea, and it is a rich source of secondary metabolites, including limonoids and protolimonoids. A collection of 36 ethanol extracts from different anatomical parts (aerial, leaves, stems, roots, fruits, and seeds) of 27 plant species, native to Cerrado from the Midwest of Brazil, were evaluated regarding their effects against Ae. aegypti larvae. Only the extract obtained from the seeds of Guarea kunthiana (Meliaceae) was shown to be active. A bioassay-guided investigation of the ethanol extract showed the larvicidal activity containing two principal constituents, the isolation and characterization of the known protolimonoids melianodiol (11) ([Table 2]) and meliantriol (12) ([Table 2]). In the evaluation of toxicity against Ae. Aegypti, only melianodiol was biologically active, with an LC50 = 28 µM. The result revealed that this natural product could be a potential candidate for the development of new biocontrol agents against Ae. aegypti larvae [71].
In a study with the hexane and ethanol extracts of 27 plant species of the Cerrado biome, the activity against Ae. aegypti larvae was tested. The species tested were prepared in five different concentrations. At 500 µg/mL, activity was observed in the ethanol extracts of A. crassiflora, Serjania lethalis, and Xylopia aromatica, and in the hexane extracts of A. crassiflora, Duguetia furfuracea, Piptocarpha rotundifolia, Casearia sylvestris var. lingua, and Cybista xantisyphilitica. These species presented larvicidal properties against the mosquito, and the authors indicate a great potential for the future isolation of active chemical compounds. The larvicidal activity was observed with a mortality of over 65% of larvae in 24 h. Different plant parts were tested, and the predominance of activity in this study was in the root and stem extracts, suggesting the presence of important secondary metabolites responsible for the activity that is biosynthesized in these parts of the plant. The authors do not provide information about the compounds responsible for the larvicidal activity, but further studies on the activity found that these extracts can contribute in controlling the Ae. aegypti vector [93].
T. avellanedae is a typical tree from Cerrado used in popular medicine and has been studied for its anti-inflammatory, antioxidant, and anticancer properties. It has been extensively studied for the effect of constituents, mainly quinones, which demonstrated potent larvicidal activity against the mosquito species Ae. aegypti. Tests with three different extracts (ethyl acetate, ethanol, and acetone) obtained from T. avellanedae wood residue indicated the presence of phenols and tannins. The larvicidal activity of acetone, ethanol, and ethyl acetate extracts against larvae of Ae. aegypti were tested and presented toxicity depending on the concentrations used. All extracts showed that activity increased with increasing exposure time. The ethanol extract, with an LC95 = 412.3 µg/mL, promoted activity for a period of 96 h, while the ethyl acetate extract had an LC95 = 319 µg/mL for a period of 48 h, and the ethanol solution reached 98.7% mortality in the first 12 h. The oviposition deterrent activity of T. avellanedae against adult Ae. aegypti females with acetone, ethyl acetate, and ethanolic extracts were more effective at 333.3 µg/mL in comparison with controls (p < 0.01). The ethyl acetate was an oviposition deterred in all concentrations tested. Reports in the literature showed that chemical composition can be similar from other studies, depending on the type of plant, plant part, and different ways of extraction. Regarding the extract of the species T. avellanedae and the presence of bioactive compounds, further studies were carried out corroborating the substantial larvicidal and oviposition repellency against Ae. aegypti, with it being a potential source to control dengue vectors [43].
Sapindus saponaria is a plant in which the bark, root, and fruit are used in folk medicine as a tranquilizer, astringent, diuretic, expectorant, tonic, blood purifier, and against cough. A study tested the morphological changes and larvicidal activity of S. saponaria collected in the Cerrado region of the Brazilian city Goiânia against Ae. aegypti larvae. In order to analyze larvicidal activity, morphological changes, and lethal mechanisms on Ae. aegypti larvae with the application of botanical insecticides, a study showed that the main action of these insecticides occurs mainly at mesenteric cells [94]. The ethanol extract fruit peel from S. saponaria was prepared in different concentrations and tested with the larvae up to 48 h. In 100 ppm, it achieved 100% mortality of Ae. aegypti larvae after 3 h, 70% in 75 ppm and 40% in 50 ppm. With 30 min of larvae exposure to the ethanol extract in 100 ppm and 75 ppm, they started to lose mobility, however, in larvae exposure to 50 ppm, the effect was observed after 2 h. There was a loss of larvicidal activity of the extract with increased storage time, indicating possible degradation of the active compounds. The morphohistological study showed damages to the mesenteric cells of Ae. aegypti. This is indicative that the chemical components present in S. saponaria can be an alternative method to the control of Ae. aegypti. The knowledge of the mechanism of action of these compounds are important for intensification of the effects and development of insecticidal products [95].
The Anacardiaceae family is characterized by the presence of flavonoids, terpenes, steroids, xanthones, phenolic, and catecholic compounds, called phenolic lipids, that usually show toxic or allergenic proprieties [96]. In the composition of this familyʼs species, there are terpenes in which the components are widely studied due to the important biological activity against Ae. aegypti. Anacardium humile leaves were collected in the Cerrado region of Campo Grande and the hexane, ethanol, and aqueous extracts and oil from the leaves were prepared in different concentrations. No toxicity was found for hexane, ethanol, and aqueous extracts against Ae. aegypti larvae, however, the essential oil of the leaves presented 100% larvae mortality in concentrations up 0.125% (LC50 = 20.9 ppm). Further studies about the pharmacologic profile of A. humile could provide more information on the favorable results for the control of Ae. aegypti [28].
A collection of 29 Cerrado species were collected in the Brazilian city Brasília, and insecticidal activity of 67 plant ethanol, hexane, dichloromethane, and hydroalcoholic extracts were evaluated at the 500 µg/mL concentration against Ae. aegypti larvae. The dichloromethane extract from Kielmeyera coriacea leaves provided activity greater than 90%, with an LC50 = 112.79 µg/mL. A few other extracts showed activity equal to or greater than 50%: hexane extracts of stem bark from Talauma ovata, root wood and leaves from Schinus terebinthifolius, and root bark from Matayba guianensis; ethanol extracts of bark and stem wood of Xylopia emarginata and leaf dichloromethane extract from S. terebinthifolius. The chemical composition of K. coriacea identified the presence of flavonoids, steroids, terpenes, and tannins in the ethanol extract [97]. Future studies may analyze the active extracts, mainly K. coriacea, that showed activity against Ae. aegypti larvae, contributing to the search for new natural compounds with larvicidal activity [98].
#
Molecular Targets for the Design of Compounds to Control Aedes aegypti
A rational strategy to address the resistance developed by Ae. aegypti against current insecticides is the design of compounds with different mechanisms of action from the ones currently in use. Rational design in the search for new drugs and active compounds is fundamental for the success in developing new products. Collaboration among researchers in the fields of ecology, molecular biology, chemistry, and medicine is a good strategy for the development of prototypes for the control of Ae. aegypti. From a natural selection point of view, it is expected that the metabolites produced by an organism have a utility for their producer, and therefore natural products are produced in order to make specific interactions with biomolecules. This is one of the reasons why natural products are the primordial source of medicinal compounds and are therefore of great interest for medicinal chemistry [99]. However, these compounds are biosynthesized and optimized for the biological system of their producer, with specific pharmacokinetic properties. Through synthetic or semisynthetic structural modifications, the chemical structure of bioactive compounds isolated from an organism can be optimized to achieve the desired bioavailability for anthropological or medicinal use. In the stages of optimization of molecular properties, the structure of the compounds can be modified with the objective of enhancing their pharmacodynamic (potency, affinity, selectivity) and pharmacokinetic properties (absorption, distribution, metabolism, excretion), and to minimize toxicity [100].
The design of active compounds is mainly based on the knowledge of the mechanisms by which bioactive molecules interact with their molecular targets [101]. Structure-based drug design by means of molecular modeling is one of the most efficient strategies used to evaluate the interaction of active compounds and its biological target, and improve pharmacodynamics [102]. This is a useful method to identify crucial intermolecular interactions in the process of molecular recognition and for the selection of hits that bind or fit to the target protein binding cavity. In the search for new compounds to be used for mosquito control, the understanding of the molecular mechanism of disease transmission by the mosquitoes is fundamental. One rational approach is to search or design compounds that interact with a biological target in order to modulate its activity and impair mosquito activity. The target is ideally a protein specific to the mosquito in order to minimize toxicity for humans and other animals. A few targets are herein described, and represented in [Fig. 1], in order to provide molecular aspects important to consider in the design of compounds for mosquito control.

The isomerase FKBP is described as the molecular target for the immunosuppressive drug tacrolimus, strongly binding in a hydrophobic pocket (13) ([Fig. 2]) and blocking the signaling pathway mediated by calcineurin [103]. The solution structure of the FKBP12 protein from Ae. aegypti (AaFKBP12, PDB ID 3UQI) was determined with 1.3 Å resolution [104]. In addition, a (3-(N-morpholino)propanesulfonic acid) buffer molecule was crystallized in the active site, which could be an interesting nucleus for designing inhibitors for AaFKBP12. Sequence similarity with HsFKBP12 is 72% and strategies should be addressed to design selective compounds to avoid toxicity to humans. The structure of AaFKBP12 might provide a useful target for the design of potential ligands to control the dengue-transmitting vector [103], [104].

The chemical communication and olfactory system of mosquitoes is also a strategic target for the development of repellents and attractants in order to control vector populations and, thus, disease transmission. OBPs are the carriers of semiochemicals that trigger signaling to activate odorant receptors [105], [106]. A series of potential ligands was evaluated to define requirements for a good fitting, and a structure-activity relationship study demonstrated that the best ligands were compounds with two aromatic rings connected by a short rigid chain [107]. Insect OBPs differ from vertebrate OBPs and have a hydrophobic cavity that encapsulate ligands and carry OBP-semiochemical complexes through the aqueous sensillar lymph to transmembrane odor receptors [91]. A study reported the structure of the major odorant-binding protein (AaegOBP1 = AaegOBP39) from Ae. aegypti describing its binding pocket, a continuous hydrophobic tunnel that involves a dimer of this protein, but no low molecular weight ligand was co-crystalized in the structure determination studies. Also, it has been proposed that binding affinity and protein conformation of the binding pocket is pH dependent. The structure of the protein was deposited in the PDB (3K1E) and is available for computational studies [105]. Also, an in vitro assay to measure affinity of ligands is described with OBP22 using N-phenyl-1-naphtylamine as the fluorescent reporter. The probe is excited at 337 nm and emission spectra can be recorded between 380 and 450 nm. The Ae. aegypti complete set of olfactory genes includes 66 putative OBPs and these could be interesting targets for mosquito control agent designs [108].
Another target is the mJHBP (PDB code 5V13) belonging to the insect OBP family and related to the D7 proteins. The JH (14) ([Fig. 3]), an insect growth regulator responsible for development, metamorphosis, reproductive development, and mating behavior, was reported to bind in the binding pocket of the N-terminal domain, and the C-terminal domain extends over the surface of the N-terminal domain and closes the entry to the binding pocket of mJHBP. The protein is selective to binding the 10R enantiomer of JH. The epoxy end of JH III is located deep in the core of the domain, and the methyl ester group is oriented toward its surface. There is a hydrogen bond between the epoxy group and the phenolic hydroxyl of Tyr-129, and the isoprenoid chain is surrounded by hydrophobic residues. Only methyl ester epoxides are able to bind to mJHBP [109]. The structural information on the binding mode of the natural substrate can be used to design substrate-mimetic ligands, a useful strategy to achieve good selectivity.

The blood feeding of the female mosquito is a key event for transmitting arboviruses. During this event, the mosquito release, in the saliva, anti-inflammatory agents and proteins to counteract these mediators and extend the period for feeding. One such protein is D7, a biogenic amine-binding salivary protein. Proteins like D7 are responsible to avoid host defensive behaviors, and therefore inhibiting them might interrupt feeding. The structure of the D7 from Ae. aegypti was determined recently, a protein that acts to neutralize two classes of inflammatory mediators. The N-terminal domain leads to the protein interior and is lined mainly with hydrophobic residues. It may accommodate bioactive lipid mediators involved in host inflammatory responses to mosquito feeding, specifically leukotrienes. A few leukotrienes (15 – 19) ([Fig. 4]) were tested in binding assays, and the highest binding affinity was observed for LTCs (LTC4, LTD4, and LTE4). The lipid chain is suggested to be the primary contact point with the protein and the peptide portion of the compounds may play a minor role in binding. Structure-activity relationships were determined for these structures and could be useful in the design of new active compounds. Additionally, norepinephrine (20) ([Fig. 4]) was detected binding to the C-terminal domain. The protein structures are deposited in the PDB codes 3DXL (unliganded structure), 3DZT (LTE4 complex), and 3DYE (norepinephrine complex). These structures could provide a suitable target with two binding pockets for computational studies and design of compounds to interrupt mosquito feeding. Inhibition of these proteins would prevent feeding and reduce the efficiency of virus transmission [110].

Another target is PK, which is responsible for catalyzing the last stage of the glycolytic pathway. It catalyzes the transphosphorylation from phosphoenolpyruvate and ADP to pyruvate and ATP. Glycolysis provides energy and intermediate precursors to other important metabolic pathways to assure limited flight performance and survival time. The activity of PK in Ae. aegypti mosquitoes has been shown to be elevated throughout embryogenesis, thus suggesting an increase of glycolysis in Ae. aegypti embryos. Thus, PK regulation becomes crucial to the metabolic needs of the mosquito. The protein AaPK1 was characterized to clarify the factors that regulate the enzyme. In order to maximize the mosquitoʼs survival time, they seek nectar for carbohydrates, and feeding sugars increase mosquito survival. Carbohydrates are necessary for blood meal digestion and reproduction. Given these factors, this study demonstrates that AaPK1 is regulated by specific sugars, amino acids, and phosphorylated sugars that are responsible for regulating important properties [111].
aaNATs are involved in the development of the exoskeleton and detoxification process in mosquitoes. aaNATs are important for mosquito survival and pose an interesting target to impair mosquito development. The structure of mosquito aaNATs are considerably different from human ones, and inhibitors of the mosquito aaNATs are expected not to interact with mammalian ones. In Ae. aegypti, there are 13 aaNATs that have been identified, showing the significance of arylakylamine acetylation in these insect species. The characterization of aaNAT structures from insects is essential for investigating the chemistry and physiology of aaNATs in mosquitoes, and it is a reference to develop new research about this family of proteins, which are essential in several insects that are vectors of diseases. Crystal structures of aaNAT2, aaNAT5b, and aaNAT7 are available in the PDB and consist of possible targets for developing new insecticides [112].
AeKAT can catalyze amino acids and keto acids that are biologically relevant. They have important activities in the central nervous system, and further investigations are needed to provide more information for their catalytic mechanism and biochemical differences in mosquitoes and humans. The crystal structure of AeKAT is available in the PDB for further studies [113]. AGTs are typical of dypteran insects such as Ae. aegypti and are catalysts for glyoxylic acid conversion to glycine. Mosquitoes are unique in possessing two AGTs, AeAGT and AeHKT, the latter being important in tryptophan metabolism for the transamination of 3-hydroxykynurenine to xanthurenic acid. AeHKT is only active on certain mosquito stages and its substract is the precursor for mosquito eye pigments. These functional adaptation and evolution aspects might be an interesting strategy to develop aminotransferase inhibitors that could be used for mosquito control. Crystal structures of AeAGT and AeHKT are available in the PDB [114].
CHDs are a class of enzymes that contribute to the interaction between DNA and nucleosomes involved in cellular processes such as replication, transcription, recombination, repair, and development. CHDs are involved in the replication of viruses, and homologs of CHDs were identified in Ae. aegypti. A significant reduction of its expression has been verified in female mosquitoes infected with Wolbachia. AeCHD7 is highly expressed during dengue infection in Ae. aegypti and seems required for DENV replication [115]. AeCHD7 could be an interesting target for the design of compounds to reduce DENV replication in mosquitoes.
Metabolic resistance to insecticides in Ae. aegypti involves alterations in the expression of a complex group of enzymes, which increases the detoxification process. These enzymes are mainly GSTs, esterases, and cytochrome P450 monooxygenases (P450). Ae. aegypti genome project revealed a set of 160 P450 genes, 49 GSTs, and 26 esterases [116].
GSTs are responsible for catalyzing the conjugation of glutathione to xenobiotic compounds in the detoxification process [117]. This enzyme was proposed to play a role in insect chemodetection by protecting the chemosensory system. Most of the GSTs comprise a super family structure of cytosolic-dimeric enzymes grouped into six classes, namely, Delta, Epsilon, Omega, Sigma, Theta, and Zeta [118]. The two major classes, Delta and Epsilon, are insect specific and are often involved with resistance [119], [120], [121]. The insecticides observed to be metabolized by GSTs include organophosphates and organochlorides [122], [123]. It was shown that DDT dehydrochlorination mediated by the Epsilon class GST confers resistance in Ae. aegypti and other mosquito species [118], [124], [125]. GST contribution to pyrethroid resistance is based in two modes of action. One is GST binding and seizure of pyrethroid, inhibiting GST activity [123], [126]. The second is through the GST protective role in oxidative stress, as the oxidative stress is a consequence of pyrethroid toxicity [127]. Generally, all GSTs adopt a very similar conserved tertiary structure, and for a functional active site, a dimeric quaternary structure is essential, as the active site is formed by amino acid residues from both subunits [128], [129]. GST inhibition could be an adjuvant strategy to reduce resistance acquired by mosquitoes.
In insects, cholesterol is required as a component of cellular membranes and as a precursor of the insect molting hormone, named ecdysone. This hormone ensures normal growth, development, and reproduction [130], [131]. However, insects lack several enzymes in the cholesterol biosynthesis pathway [132], which make them unable to synthesize cholesterol de novo [130], [132]. Thus, it is essential that the insects obtain cholesterol through a dietary source, making cholesterol uptake and transport potential targets for the development of new mosquito larvicides [133]. In insects, SCP-2 is involved in the absorption of lipid droplets to the cytoplasmic membrane in the body fat or intracellular cholesterol transportation from the luminal side to the basal side of the midgut epithelium [134], [135], [136]. As cholesterol is highly hydrophobic, carrier proteins mediate its delivery by shielding its hydrophobic moiety from the aqueous environment of the cytoplasm [137], [138], [139]. SCP-2 belongs to the SCP-2 gene family, which contains a sterol-binding domain located at the C-terminus, identified in vertebrates, insects, plants, yeast, bacteria, and fungi [140], [141]. AeSCP-2 is expressed mainly at the cytoplasm [142], [143], [144] and has been shown to aid in the uptake of cholesterol in mosquito cells, mosquito development, and reproduction [134], [145]. AeSCP-2 is mainly expressed throughout the feeding larval stages in midgut tissue, the main site of cholesterol absorption [142]. AeSCP-2 knockdown resulted in a reduction of cholesterol uptake in larvae and high larval mortality. In adults, it caused reduced, inefficient cholesterol uptake after blood meals and a fecundity decrease [145]. However, it had little effect on palmitic acid uptake [145]. Studies also demonstrated that DENV replication in human host cells depends on both de novo cholesterol biosynthesis and intracellular transport. When SCP-2 function was inhibited, the production of DENV viral particles was impaired, probably because the SCP-2 mediates the cholesterol trafficking pathway, which is critical for DENV production. AeSCP-2 is different from vertebrate SCP-2, with their hydrophobic moieties oriented at the opposite ends of the protein. Moreover, AeSCP-2 is essential for mosquito survival and development, whereas vertebrate SCP-2 is not vital for their survival and fertility genomic data have shown that Ae. aegypti has four genes encoding single SCP-2 domain proteins [141], [144]. AeSCP-2 and AeSCP-2-like proteins have very similar temporal and spatial expression profiles with variable affinity to different lipids [141], [142]. It was shown that AeSCP-2 contributes to the uptake of cholesterol from a blood meal in female mosquitoes and AeSCP-2-like protein affects the uptake of free fatty acids [145]. This carrier protein could be used as a target for drug design in order to selectively control mosquito populations.
#
Conclusions
The impact caused by arboviruses diseases on the health systems of tropical countries is huge. Although arboviruses cause a great health impact in Brazil, there are only a few studies of the Cerrado extensive biodiversity for new insecticides to control their vector. Vector control is a useful strategy to control transmission of these diseases, with the mosquito Ae. aegypti being the major target in this context. Natural products from the Brazilian biome Cerrado are highlighted as a valuable source of agents with potential to be sustainably explored by our health systems and industries for the development of products for mosquito control. The development of such products requires optimization of pharmacodynamic and pharmacokinetic properties that might be tackled by a medicinal chemistry approach. The study of the mechanisms of action of active compounds and their biological targets provide important knowledge in this task. An extensive search of targets was herein presented as insights to promote advances in the highly difficult task of developing products to control Ae. aegypti.
#
Contributorsʼ Statement
Designed the work: M. Valli, A. D. Andricopulo, L. S. Espindola, V. S. Bolzani; collected and analysed data: M. Valli, L. C. V. Atanázio, G. C. Monteiro, R. R. Coelho, D. P. Demarque; supervised and discussed collection of data: A. D. Andricopulo, L. S. Espindola, V. S. Bolzani; drafted the manuscript: M. Valli, L. C. V. Atanázio, G. C. Monteiro, R. R. Coelho, D. P. Demarque. All authors revised the manuscript.
#
#
Conflict of Interest
The authors declare that they have no conflict of interest.
Acknowledgements
The authors acknowledge the Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP), grants #2013/07600-3 (CIBFar-CEPID), and #2014/50926-0 (INCT BioNat CNPq/FAPESP), and the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for grant support and research fellowships. Fundação de Apoio a Pesquisa do Distrito Federal (FAPDF), grants #0193.001482/2016. ArboControl Brasil Project – Process TED 74/2016 and TED 42/2017 (Fundo Nacional de Saúde/Universidade de Brasília) for funding and post-doctoral fellowships (M. V., G. C. M., R. C., and D. P. D.). M. V. also acknowledges the scholarship FAPESP #2019/05967-3.
-
References
- 1 Newman DJ, Cragg GM. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J Nat Prod 2020; 83: 770-803
- 2 Saldivar-Gonzalez FI, Valli M, Andricopulo AD, da Silva Bolzani V, Medina-Franco JL. Chemical space and diversity of the NuBBE database: A chemoinformatic characterization. J Chem Inf Model 2019; 59: 74-85
- 3 Myers N, Mittermeier RA, Mittermeier CG, da Fonseca GA, Kent J. Biodiversity hotspots for conservation priorities. Nature 2000; 403: 853-858
- 4 Eiten G. Cerrado: Caracterização, Ocupação e Perspectivas. In: Pinto MN. ed. Vegetação do Cerrado. Brasília: Editora da Universidade de Brasília; 1994: 17-73
- 5 Simon MF, Grether R, de Queiroz LP, Skema C, Pennington RT, Hughes CE. Recent assembly of the Cerrado, a neotropical plant diversity hotspot, by in situ evolution of adaptations to fire. Proc Natl Acad Sci U S A 2009; 106: 20359-20364
- 6 Machado R, Neto M, Pereira P, Caldas E, Gonçalves D, Santos N, Tabor K, Steininger M. Estimativas de perda da área do Cerrado brasileiro. Brasília, DF: Conservação Internacional; 2004
- 7 Flausino O, Valli M, Bolzani V. Biodiversidade brasileira: uma fonte potencial de agentes terapêuticos ainda inexplorada. In: Yunes R, Cechinel Filho V. eds. Química de produtos naturais: novos fármacos e a moderna farmacognosia. 3rd ed.. Itajaí: Univali; 2012: 353-383
- 8 Pilon AC, Valli M, Dametto AC, Pinto MEF, Freire RT, Castro-Gamboa I, Andricopulo AD, Bolzani VS. NuBBEDB: an updated database to uncover chemical and biological information from Brazilian biodiversity. Sci Rep 2017; 7: 7215
- 9 Pierson TC, Diamond MS. The continued threat of emerging flaviviruses. Nat Microbiol 2020; 5: 796-812
- 10 Sinka ME, Bangs MJ, Manguin S, Rubio-Palis Y, Chareonviriyaphap T, Coetzee M, Mbogo CM, Hemingway J, Patil AP, Temperley WH, Gething PW, Kabaria CW, Burkot TR, Harbach RE, Hay SI. A global map of dominant malaria vectors. Parasit Vectors 2012; 5: 69
- 11 Dunphy BM, Kovach KB, Gehrke EJ, Field EN, Rowley WA, Bartholomay LC, Smith RC. Long-term surveillance defines spatial and temporal patterns implicating Culex tarsalis as the primary vector of West Nile virus. Sci Rep 2019; 9: 6637
- 12 Leparc-Goffart I, Nougairede A, Cassadou S, Prat C, de Lamballerie X. Chikungunya in the Americas. Lancet 2014; 383: 514
- 13 Kraemer MU, Sinka ME, Duda KA, Mylne AQ, Shearer FM, Barker CM, Moore CG, Carvalho RG, Coelho GE, Van Bortel W, Hendrickx G, Schaffner F, Elyazar IR, Teng HJ, Brady OJ, Messina JP, Pigott DM, Scott TW, Smith DL, Wint GR, Golding N, Hay SI. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus . Elife 2015; 4: e08347
- 14 Wilder-Smith A, Ooi EE, Horstick O, Wills B. Dengue. Lancet 2019; 393: 350-363
- 15 WHO. Dengue and severe dengue. Accessed May 30, 2020 at: https://www.who.int/news-room/fact-sheets/detail/dengue-and-severe-dengue
- 16 Brady OJ, Gething PW, Bhatt S, Messina JP, Brownstein JS, Hoen AG, Moyes CL, Farlow AW, Scott TW, Hay SI. Refining the global spatial limits of dengue virus transmission by evidence-based consensus. PLoS Negl Trop Dis 2012; 6: e1760
- 17 Bhatt S, Gething PW, Brady OJ, Messina JP, Farlow AW, Moyes CL, Drake JM, Brownstein JS, Hoen AG, Sankoh O, Myers MF, George DB, Jaenisch T, Wint GR, Simmons CP, Scott TW, Farrar JJ, Hay SI. The global distribution and burden of dengue. Nature 2013; 496: 504-507
- 18 PAHO. Casos reportados de dengue en las Américas. Accessed May 30, 2020 at: https://www.paho.org/data/index.php/es/temas/indicadores-dengue/dengue-nacional/9-dengue-pais-ano.html
- 19 WHO. Zika virus. Accessed May 30, 2020 at: https://www.who.int/en/news-room/fact-sheets/detail/zika-virus
- 20 Brasil P, Pereira JP, Moreira ME, Ribeiro Nogueira RM, Damasceno L, Wakimoto M, Rabello RS, Valderramos SG, Halai UA, Salles TS, Zin AA, Horovitz D, Daltro P, Boechat M, Raja Gabaglia C, Carvalho de Sequeira P, Pilotto JH, Medialdea-Carrera R, Cotrim da Cunha D, Abreu de Carvalho LM, Pone M, Machado Siqueira A, Calvet GA, Rodrigues Baiao AE, Neves ES, Nassar de Carvalho PR, Hasue RH, Marschik PB, Einspieler C, Janzen C, Cherry JD, Bispo de Filippis AM, Nielsen-Saines K. Zika virus infection in pregnant women in Rio de Janeiro. N Engl J Med 2016; 375: 2321-2334
- 21 WHO. Chikungunya. Accessed May 30, 2020 at: https://www.who.int/news-room/fact-sheets/detail/chikungunya
- 22 Guy B, Briand O, Lang J, Saville M, Jackson N. Development of the Sanofi Pasteur tetravalent dengue vaccine: One more step forward. Vaccine 2015; 33: 7100-7111
- 23 Guy B, Jackson N. Dengue vaccine: hypotheses to understand CYD-TDV-induced protection. Nat Rev Microbiol 2016; 14: 45-54
- 24 WHO. Questions and Answers on Dengue Vaccines. Accessed May 30, 2020 at: https://www.who.int/immunization/research/development/dengue_q_and_a/en/
- 25 WHO. Neglected tropical diseases. Accessed May 30, 2020 at: https://www.who.int/neglected_diseases/en/
- 26 Barbosa PBB, Oliveira JM, Chagas JM, Rabelo LMA, Medeiros GF, Giodani RB, Silva EA, Uchôa AF, Ximenes MFF. Evaluation of seed extracts from plants found in the Caatinga biome for the control of Aedes aegypti . Parasitol Res 2014; 113: 3565-3580
- 27 Ministério da Saúde. Fundação Nacional de Saúde. Manual de vigilância epidemiológica de febre amarela. Brasília, DF: Ministério da Saúde: Fundação Nacional de Saúde; 2004. Accessed December 15 2020 at: https://bvsms.saude.gov.br/bvs/publicacoes/manual_vigilancia_epid_febre_amarela.pdf
- 28 Porto KR, Roel AR, Silva MM, Coelho RM, Scheleder EJ, Jeller AH. Atividade larvicida do óleo de Anacardium humile Saint Hill sobre Aedes aegypti (Linnaeus, 1762) (Diptera, Culicidae). Rev Bras Med Trop 2008; 41: 586-589
- 29 de Mendonca FA, da Silva KF, dos Santos KK, Ribeiro Júnior KA, SantʼAna AE. Activities of some Brazilian plants against larvae of the mosquito Aedes aegypti . Fitoterapia 2005; 76: 629-636
- 30 de Carvalho GHF, de Andrade MA, de Araujo CN, Santos ML, de Castro NA, Charneau S, Monnerat R, de Santana JM, Bastos IMD. Larvicidal and pupicidal activities of eco-friendly phenolic lipid products from Anacardium occidentale nutshell against arbovirus vectors. Environ Sci Pollut Res Int 2019; 26: 5514-5523
- 31 Souza TM, Farias DF, Soares BM, Viana MP, Lima GPG, Machado LKA, Morais SM, Carvalho AFU. Toxicity of Brazilian plant seed extracts to two strains of Aedes aegypti (Diptera: Culicidae) and nontarget animals. J Med Entomol 2011; 48: 846-851
- 32 Napoleao TH, Pontual EV, de Albuquerque Lima T, de Lima Santos ND, Sá RA, Coelho LC, do Amaral Ferraz Navarro DM, Paiva PM. Effect of Myracrodruon urundeuva leaf lectin on survival and digestive enzymes of Aedes aegypti larvae. Parasitol Res 2012; 110: 609-616
- 33 Silva AG, Almeida DL, Ronchi SN, Bento AC, Scherer R, Ramos AC, Cruz ZM. The essential oil of Brazilian pepper, Schinus terebinthifolia Raddi in larval control of Stegomyia aegypti (Linnaeus, 1762). Parasit Vectors 2010; 3: 79
- 34 Santos CCS, Araujo SS, Santos ALLM, Almeida ECV, Dias AS, Damascena NP, Santos DM, Santos MIS, Júnior KALR, Pereira CKB, Lima ACB, Shan AYKV, Santʼana AEG, Estevam CS, Araujo BS. Evaluation of the toxicity and molluscicidal and larvicidal activities of Schinopsis brasiliensis stem bark extract and its fractions. Rev Bras Farmacogn 2014; 24: 298-303
- 35 de Omena MC, Navarro DMAF, de Paula JE, Luna JS, de Lima MRF, SantʼAna AEG. Larvicidal activities against Aedes aegypti of some Brazilian medicinal plants. Bioresour Technol 2007; 98: 2549-2556
- 36 Grzybowski A, Tiboni M, Silva MA, Chitolina RF, Passos M, Fontana JD. Synergistic larvicidal effect and morphological alterations induced by ethanolic extracts of Annona muricata and Piper nigrum against the dengue fever vector Aedes aegypti . Pest Manag Sci 2013; 69: 589-601
- 37 Rodrigues AM, Silva AAS, Pinto CCC, Lima Dos Santos D, Carneiro de Freitas JC, Martins VEP, Maia de Morais S. Larvicidal and enzymatic inhibition effects of Annona muricata seed extract and main constituent annonacin against Aedes aegypti and Aedes albopictus (Diptera: Culicidae). Pharmaceuticals (Basel) 2019; 12: 112
- 38 Simas NK, Dellamora EDL, Schripsema J, Lage CLS, de Oliveira AM, Wessjohann L, Porzel A, Kuster RM. Acetylenic 2-phenylethylamides and new isobutylamides from Acmella oleracea (L.) R. K. Jansen, a Brazilian spice with larvicidal activity on Aedes aegypti . Phytochem Lett 2013; 6: 67-72
- 39 Lavor PL, Santiago GMP, Gois RWD, de Sousa LM, Bezerra GD, Romero NR, Arriaga AMC, Lemos TLG, Alves PB, Gomes PCS. Larvicidal activity against Aedes aegypti of essential oils from northeast Brazil. Nat Prod Commun 2012; 7: 1391-1392
- 40 Botas GDS, Cruz RAS, de Almeida FB, Duarte JL, Araujo RS, Souto RNP, Ferreira R, Carvalho JCT, Santos MG, Rocha L, Pereira VLP, Fernandes CP. Baccharis reticularia DC. and limonene nanoemulsions: Promising larvicidal agents for Aedes aegypti (Diptera: Culicidae) control. Molecules 2017; 22: 1990
- 41 Marques MM, Morais SM, Vieira IG, Vieira MG, Raquel A, Silva A, De Almeida RR, Guedes MI. Larvicidal activity of Tagetes erecta against Aedes aegypti . J Am Mosq Control Assoc 2011; 27: 156-158
- 42 Garcez WS, Garcez FR, da Silva LMGE, Hamerski L. Larvicidal activity against Aedes aegypti of some plants native to the West-Central region of Brazil. Bioresour Technol 2009; 100: 6647-6650
- 43 Borges JCM, Silva EAP e, de Barros TCA, Soares IM, Ascêncio SD, Fidelis RR, de Souza Aguiar RW. Chemical composition, oviposition deterrent and larvicidal activities of the wood extracts of Tabebuia avellanedae from the Cerrado of Brazil. J Med Plant Res 2018; 12: 404-414
- 44 Costa JGM, Pessoa ODL, Menezes EA, Santiago GMP, Lemos TLG. Composition and larvicidal activity of essential oils from heartwood of Auxemma glazioviana Taub. (Boraginaceae). Flavour Frag J 2004; 19: 529-531
- 45 Santos RP, Nunes EP, Nascimento RF, Santiago GMP, Menezes GHA, Silveira ER, Pessoa ODL. Chemical composition and larvicidal activity of the essential oils of Cordia leucomalloides and Cordia curassavica from the northeast of Brazil. J Brazil Chem Soc 2006; 17: 1027-1030
- 46 de Menezes JESA, Lemos TLG, Silveira ER, Pessoa ODL, Santiago GMP. Chemical composition and larvicidal activity of the essential oil from leaves of Cordia globosa (Jacq.) HBK from northeastern Brazil. J Essent Oil Res 2006; 18: 253-255
- 47 da Silva RC, Milet-Pinheiro P, Bezerra da Silva PC, da Silva AG, da Silva MV, Navarro DM, da Silva NH. (E)-caryophyllene and alpha-humulene: Aedes aegypti oviposition deterrents elucidated by gas chromatography-electrophysiological assay of Commiphora leptophloeos leaf oil. PLoS One 2015; 10: e0144586
- 48 Oliveira PV, Ferreira JC, Moura FS, Lima GS, de Oliveira FM, Oliveira PE, Conserva LM, Giulietti AM, Lemos RP. Larvicidal activity of 94 extracts from ten plant species of northeastern of Brazil against Aedes aegypti L. (Diptera: Culicidae). Parasitol Res 2010; 107: 403-407
- 49 de Lima GP, de Souza TM, de Paula Freire G, Farias DF, Cunha AP, Ricardo NM, de Morais SM, Carvalho AF. Further insecticidal activities of essential oils from Lippia sidoides and Croton species against Aedes aegypti L. Parasitol Res 2013; 112: 1953-1958
- 50 Morais SM, Cavalcanti ESB, Bertini LM, Oliveira CLL, Rodrigues JRB, Cardoso JHL. Larvicidal activity of essential oils from Brazilian Croton species against Aedes aegypti L. J Am Mosq Control Assoc 2006; 22: 161-164
- 51 Doria GA, Silva WJ, Carvalho GA, Alves PB, Cavalcanti SC. A study of the larvicidal activity of two Croton species from northeastern Brazil against Aedes aegypti . Pharm Biol 2010; 48: 615-620
- 52 Santos GK, Dutra KA, Lira CS, Lima BN, Napoleao TH, Paiva PM, Maranhao CA, Brandao SS, Navarro DM. Effects of Croton rhamnifolioides essential oil on Aedes aegypti oviposition, larval toxicity and trypsin activity. Molecules 2014; 19: 16573-16587
- 53 Torres MC, Assuncao JC, Santiago GM, Andrade-Neto M, Silveira ER, Costa-Lotufo LV, Bezerra DP, Marinho Filho JD, Viana FA, Pessoa OD. Larvicidal and nematicidal activities of the leaf essential oil of Croton regelianus . Chem Biodivers 2008; 5: 2724-2728
- 54 Carvalho Kda S, Silva SL e, de Souza IA, Gualberto SA, da Cruz RC, Dos Santos FR, de Carvalho MG. Toxicological evaluation of essential oil from the leaves of Croton tetradenius (Euphorbiaceae) on Aedes aegypti and Mus musculus . Parasitol Res 2016; 115: 3441-3448
- 55 Farias DF, Cavalheiro MG, Viana MP, Queiroz VA, Rocha-Bezerra LCB, Vasconcelos IM, Morais SM, Carvalho AFU. Water extracts of Brazilian leguminous seeds as rich sources of larvicidal compounds against Aedes aegypti L. An Acad Bras Ciênc 2010; 82: 585-594
- 56 Trindade FTT, Stabeli RG, Pereira AA, Facundo VA, Silva ADE. Copaifera multijuga ethanolic extracts, oil-resin, and its derivatives display larvicidal activity against Anopheles darlingi and Aedes aegypti (Diptera: Culicidae). Rev Bras Farmacogn 2013; 23: 464-470
- 57 Dalarmi L, da Silva CB, Ocampos FMM, Burci LM, do Nascimento KF, de Jesus C, Dias JdFG, Miguel MD, Miguel OG, Zanin SMW. Larvicidal activity of Dalbergia brasiliensis (Fabaceae – Papilionoideae) on Aedes aegypti . Afr J Pharm Pharmaco 2018; 9: 881-885
- 58 Gusmao DS, Pascoa V, Mathias L, Curcino Vieira IJ, Braz-Filho R, Alves Lemos FJ. Derris (Lonchocarpus) urucu (Leguminosae) extract modifies the peritrophic matrix structure of Aedes aegypti (Diptera: Culicidae). Mem Inst Oswaldo Cruz 2002; 97: 371-375
- 59 Aguiar JC, Santiago GM, Lavor PL, Veras HN, Ferreira YS, Lima MA, Arriaga AM, Lemos TL, Lima JQ, de Jesus HC, Alves PB, Braz-Filho R. Chemical constituents and larvicidal activity of Hymenaea courbaril fruit peel. Nat Prod Commun 2010; 5: 1977-1980
- 60 Porto KRA, Motti PR, Yano M, Roel AR, Cardoso CAL, Matias R. Screening of plant extracts and fractions on Aedes aegypti larvae found in the state of Mato Grosso do Sul (linnaeus, 1762) (culicidae). An Acad Bras Ciênc 2017; 8: 895-906
- 61 Arriaga AM, Lima JQ, Vasconcelos JN e, de Oliveira MC, Lemos TL, Fonseca AM, Malcher GT, Santiago GM, Mafezoli J, Braz-Filho R. Antioxidant and larvicidal activities of Tephrosia egregia Sandw against Aedes aegypti . Nat Prod Commun 2009; 4: 529-530
- 62 Costa JGM, Rodrigues FFG, Angélico EC, Silva MR, Mota ML, Santos NKA, Cardoso ALH, Lemos TLG. Estudo químico-biológico dos óleos essenciais de Hyptis martiusii, Lippia sidoides e Syzigium aromaticum frente às larvas do Aedes aegypti . Rev Bras Farmacogn 2005; 15: 304-309
- 63 Silva WJ, Doria GA, Maia RT, Nunes RS, Carvalho GA, Blank AF, Alves PB, Marcal RM, Cavalcanti SC. Effects of essential oils on Aedes aegypti larvae: alternatives to environmentally safe insecticides. Bioresour Technol 2008; 99: 3251-3255
- 64 Ricarte LP, Bezerra GP, Romero NR, Silva HCD, Lemos TLG, Arriaga AMC, Alves PB, Santos MBD, Militao GCG, Silva TDS, Braz-Filho R, Santiago GMP. Chemical composition and biological activities of the essential oils from Vitex-agnus castus, Ocimum campechianum and Ocimum carnosum . An Acad Bras Cienc 2020; 92: e20180569
- 65 Azevedo SG, Mar JM, da Silva LS, Franca LP, Machado MB, Tadei WP, Bezerra JD, dos Santos AL, Sanches EA. Bioactivity of Licaria puchury-major essential oil against Aedes aegypti, Tetranychus urticae and Cerataphis lataniae . Rec Nat Prod 2018; 12: 229-239
- 66 da Silva HHG, Geris R, Rodrigues E, Rocha C, da Silva IG. Larvicidal activity of oil-resin fractions from the Brazilian medicinal plant Copaifera reticulata Ducke (Leguminosae-Caesalpinoideae) against Aedes aegypti (Diptera, Culicidae). Rev Soc Bras Med Trop 2007; 40: 264-267
- 67 Gois RWD, de Sousa LM, Lemos TLG, Arriaga AMC, Andrade-Neto M, Santiago GMP, Ferreira YS, Alves PB, de Jesus HCR. Chemical composition and larvicidal effects of essential oil from Bauhinia acuruana (Moric) againstAedes aegypti. . J Essent Oil Res 2011; 23: 59-62
- 68 dos Santos EA, de Carvalho CM, Costa ALS, Conceicao AS, Moura FDP, Santana AEG. Bioactivity evaluation of plant extracts used in indigenous medicine against the snail, Biomphalaria glabrata, and the larvae of Aedes aegypti . Evid Based Complement Alternat 2012; 2012: 846583
- 69 Ferreira MDL, Fernandes DA, Nunes FC, Teles YCF, Rolim YM, da Silva CM, de Albuquerque JBL, Agra MF, de Souza MFV. Phytochemical study of Waltheria viscosissima and evaluation of its larvicidal activity against Aedes aegypti . Rev Bras Farmacogn 2019; 29: 582-590
- 70 Ferreira MR, Santiago RR, de Souza TP, Egito ES, Oliveira EE, Soares LA. Development and evaluation of emulsions from Carapa guianensis (Andiroba) oil. AAPS PharmSciTech 2010; 11: 1383-1390
- 71 Sarmento UC, Miguita CH, Almeida LH, Gaban CR, Silva LM, Souza AS, Garcez WS, Garcez FR. Larvicidal efficacies of plants from Midwestern Brazil: melianodiol from Guarea kunthiana as a potential biopesticide against Aedes aegypti . Mem Inst Oswaldo Cruz 2016; 111: 469-474
- 72 Magalhaes LAM, Lima MD, Marques MOM, Facanali R, Pinto ACD, Tadei WP. Chemical composition and larvicidal activity against Aedes aegypti larvae of essential oils from four Guarea species. Molecules 2010; 15: 5734-5741
- 73 da Silva AG, Alves RCC, Filho CMB, Bezerra-Silva PC, dos Santos LMM, Foglio MA, Navarro DMDF, da Silva MV, Correia MTD. Chemical composition and larvicidal activity of the essential oil from leaves of Eugenia brejoensis Mazine (Myrtaceae). J Essent Oil Bear Pl 2015; 18: 1441-1447
- 74 Neves ID, Rezende SRD, Kirk JM, Pontes EG, De Carvalho MG. Composition and larvicidal activity of essential oil of Eugenia candolleana DC. (MYRTACEAE) against Aedes aegypti . Rev Virtual Quim 2017; 9: 2305-2315
- 75 Dias CN, Alves LP, Rodrigues KA, Brito MC, Rosa Cdos S, do Amaral FM, Monteiro Odos S, Andrade EH, Maia JG, Moraes DF. Chemical composition and larvicidal activity of essential oils extracted from Brazilian legal amazon plants against Aedes aegypti L. (Diptera: Culicidae). Evid Based Complement Alternat Med 2015; 2015: 490765
- 76 Rosa CS, Veras KS, Silva PR, Lopes Neto JJ, Cardoso HLM, Alves LPL, Brito MCA, Amaral FMM, Maia JGS, Monteiro OS, Moraes DFC. Composição química e toxicidade frente Aedes aegypti L. e Artemia salina Leach do óleo essencial das folhas de Myrcia sylvatica (G. Mey.) DC. Rev Bras Pl Med 2016; 18: 19-26
- 77 Mendes LA, Martins GF, Valbon WR, de Souza TD, Menini L, Ferreira A, Ferreira MFD. Larvicidal effect of essential oils from Brazilian cultivars of guava on Aedes aegypti L. Ind Crop Prod 2017; 108: 684-689
- 78 Kanis LA, Antonio RD, Antunes EP, Prophiro JS, da Silva OS. Larvicidal effect of dried leaf extracts from Pinus caribaea against Aedes aegypti (Linnaeus, 1762) (Diptera: Culicidae). Rev Soc Bras Med Trop 2009; 42: 373-376
- 79 Oliveira GL, Cardoso SK, Lara CR, Vieira TM, Guimaraes EF, Figueiredo LS, Martins ER, Moreira DL, Kaplan MA. Chemical study and larvicidal activity against Aedes aegypti of essential oil of Piper aduncum L. (Piperaceae). An Acad Bras Cienc 2013; 85: 1227-1234
- 80 da Silva MF, Bezerra-Silva PC, de Lira CS, de Lima Albuquerque BN, Agra Neto AC, Pontual EV, Maciel JR, Paiva PM, Navarro DM. Composition and biological activities of the essential oil of Piper corcovadensis (Miq.) C. DC (Piperaceae). Exp Parasitol 2016; 165: 64-70
- 81 do Nascimento JC, David JM, Barbosa LC, de Paula VF, Demuner AJ, David JP, Conserva LM, Ferreira JC, Guimaraes EF. Larvicidal activities and chemical composition of essential oils from Piper klotzschianum (Kunth) C. DC. (Piperaceae). Pest Manag Sci 2013; 69: 1267-1271
- 82 Autran ES, Neves IA, da Silva CSB, Santos GKN, da Camara CAG, Navarro DMAF. Chemical composition, oviposition deterrent and larvicidal activities against Aedes aegypti of essential oils from Piper marginatum Jacq. (Piperaceae). Bioresour Technol 2009; 100: 2284-2288
- 83 Marques AM, Velozo LS, Carvalho MA, Serdeiro MT, Honorio NA, Kaplan MAC, Maleck M. Larvicidal activity of Ottonia anisum metabolites against Aedes aegypti: A potential natural alternative source for mosquito vector control in Brazil. J Vector Dis 2017; 54: 61-68
- 84 Cavalcanti ESB, de Morais SM, Lima MAA, Santana EWP. Larvicidal activity of essential oils from Brazilian plants against Aedes aegypti L. Mem Inst Oswaldo Cruz 2004; 99: 541-544
- 85 Arriaga AMC, Rodrigues FEA, Lemos TLG, de Oliveira MDF, Lima JQ, Santiago GMP, Braz-Filho R, Matezoli J. Composition and larvicidal activity of essential oil from Stemodia maritima L. Nat Prod Commun 2007; 2: 1237-1239
- 86 Ferreira RMDA, Dʼhaveloose NP, Cruz RAS, Araújo RS, Carvalho JCT, Rocha L, Fernandes LP, Da Costa TS, Fernandes CP, Souto RNP. Nano-emulsification enhances the larvicidal potential of the essential oil of Siparuna guianensis (Laurales: Siparunaceae) against Aedes (Stegomyia) aegypti (Diptera: Culicidae). J Med Entomol 2020; 57: 788-796
- 87 Avelar Amado P, Fonseca Castro AH, Nunes Alves S, Brentan Silva D, Alexandre Carollo C. Alves Rodrigues Dos Santos Lima L. Phenolic compounds: antioxidant and larvicidal potential of Smilax brasiliensis Sprengel leaves. Nat Prod Res 2019; 17: 2545-2553
- 88 Santiago GMP, Lemos TLG, Pessoa ODL, Arriaga AMC, Matos FJA, Lima MAS, Santos HS, Lima DCL, Barbosa FG, Luciano JHS, Silveira ER, de Menezes GHA. Larvicidal activity against Aedes aegypti L. (Diptera: Culicidae) of essential oils of Lippia species from Brazil. Nat Prod Commun 2006; 1: 573-576
- 89 Nascimento AMD, Maia TDS, Soares TES, Menezes LRA, Scher R, Costa EV, Cavalcanti SCH, La Corte R. Repellency and larvicidal activity of essential oils from Xylopia laevigata, Xylopia frutescens, Lippia pedunculosa, and their individual compounds against Aedes aegypti Linnaeus. Neotrop Entomol 2017; 46: 223-230
- 90 Costa M, Pereira M, Oliveira S, Souza P, Dallʼoglio E, Alves T. Anonáceas provocam mortalidade em larvas de Aedes aegypti (Linnaeus, 1762) (Dipteria: Culidae). Rev Bras Biocienc 2013; 11: 184-190
- 91 Costa MS, Santana AE, Oliveira LL, Zanuncio JC, Serrao JE. Toxicity of squamocin on Aedes aegypti larvae, its predators and human cells. Pest Manag Sci 2017; 73: 636-640
- 92 De Sousa FDM, Grossi SM, Monteiro GC, Demarque DP, Espindola LS. Dereplication and isolation of larvicidal compounds from annonaceae species against Aedes aegypti . Rev Bras Farmacogn 2020; 30: 123-126
- 93 Rodrigues AMS, De Paula JE, Degallier N, Molez JE, Espindola LS. Larvicidal activity of some Cerrado plant extracts against Aedes aegypti . J Am Mosq Control Assoc 2006; 22: 314-317
- 94 Borges RA, Cavasin GM, Silva IG, Arruda W, Oliveira ESF, Silva HHG, Martins F. Mortalidade e alterações morfológicas provocadas pela ação inibidora do diflubenzuron na ecdise de larvas de Aedes aegypti (Diptera, Culicidae). Rev Patol Trop 2004; 33: 91-105
- 95 Barreto CF, Cavasin GM, Silva HHG, Silva IG. Estudo das alterações morfo-histológicas em larvas de Aedes aegypti (Diptera, Culicidae) submetidas ao extrato bruto etanólico de Sapindus saponaria Lin (Sapindaceae). Rev Patol Trop 2006; 35: 37-57
- 96 Correia SJ, David JP, David JM. Secundary metabolites from species of Anacardiaceae. Quim Nova 2006; 29: 1287-1300
- 97 Audi EA, Otobone F, Martins JV, Cortez DA. Preliminary evaluation of Kielmeyera coriacea leaves extract on the central nervous system. Fitoterapia 2002; 73: 517-519
- 98 Coelho AAM, Paula JE, Espindola LS. Atividade larvicida de extratos vegetais sobre Aedes aegypti (L.) (Diptera: Culicidae), em condições de laboratório. Rev Entom Bras 2009; 4: 1-6
- 99 Ma DL, Chan DSH, Leung CH. Molecular docking for virtual screening of natural product databases. Chem Sci 2011; 2: 1656-1665
- 100 Andricopulo AD, Montanari CA. Structure-activity relationships for the design of small-molecule inhibitors. Mini Rev Med Chem 2005; 5: 585-593
- 101 Ferreira LG, Andricopulo AD. From protein structure to small-molecules: recent advances and applications to fragment-based drug discovery. Curr Top Med Chem 2017; 17: 2260-2270
- 102 Ferreira LG, Oliva G, Andricopulo AD. From medicinal chemistry to human health: Current approaches to drug discovery for cancer and neglected tropical diseases. An Acad Bras Cienc 2018; 90: 645-661
- 103 Chakraborty G, Shin J, Nguyen QT, Harikishore A, Baek K, Yoon HS. Solution structure of FK506-binding protein 12 from Aedes aegypti . Proteins 2012; 80: 2476-2481
- 104 Rajan S, Saw KQ, Nguyen QT, Baek K, Yoon HS. High-resolution crystal structure of FKBP12 from Aedes aegypti . Protein Sci 2012; 21: 1080-1084
- 105 Leite NR, Krogh R, Xu W, Ishida Y, Iulek J, Leal WS, Oliva G. Structure of an odorant-binding protein from the mosquito Aedes aegypti suggests a binding pocket covered by a pH-sensitive “Lid”. PLoS One 2009; 4: e8006
- 106 Leal WS. Odorant reception in insects: roles of receptors, binding proteins, and degrading enzymes. Annu Rev Entomol 2013; 58: 373-391
- 107 Li S, Picimbon JF, Ji S, Kan Y, Chuanling Q, Zhou JJ, Pelosi P. Multiple functions of an odorant-binding protein in the mosquito Aedes aegypti . Biochem Biophys Res Commun 2008; 372: 464-468
- 108 Bohbot J, Pitts RJ, Kwon HW, Rutzler M, Robertson HM, Zwiebel LJ. Molecular characterization of the Aedes aegypti odorant receptor gene family. Insect Mol Biol 2007; 16: 525-537
- 109 Kim IH, Pham V, Jablonka W, Goodman WG, Ribeiro JMC, Andersen JF. A mosquito hemolymph odorant-binding protein family member specifically binds juvenile hormone. J Biol Chem 2017; 292: 15329-15339
- 110 Calvo E, Mans BJ, Ribeiro JM, Andersen JF. Multifunctionality and mechanism of ligand binding in a mosquito antiinflammatory protein. Proc Natl Acad Sci U S A 2009; 106: 3728-3733
- 111 Petchampai N, Murillo-Solano C, Isoe J, Pizarro JC, Scaraffia PY. Distinctive regulatory properties of pyruvate kinase 1 from Aedes aegypti mosquitoes. Insect Biochem Mol Biol 2019; 104: 82-90
- 112 Han Q, Robinson H, Ding H, Christensen BM, Li J. Evolution of insect arylalkylamine N-acetyltransferases: structural evidence from the yellow fever mosquito, Aedes aegypti . Proc Natl Acad Sci U S A 2012; 109: 11669-11674
- 113 Han Q, Gao YG, Robinson H, Ding H, Wilson S, Li J. Crystal structures of Aedes aegypti kynurenine aminotransferase. FEBS J 2005; 272: 2198-2206
- 114 Han Q, Robinson H, Gao YG, Vogelaar N, Wilson SR, Rizzi M, Li J. Crystal structures of Aedes aegypti alanine glyoxylate aminotransferase. J Biol Chem 2006; 281: 37175-37182
- 115 Asad S, Hall-Mendelin S, Asgari S. Downregulation of Aedes aegypti chromodomain helicase DNA binding protein 7/Kismet by Wolbachia and its effect on dengue virus replication. Sci Rep 2016; 6: 36850
- 116 Strode C, Wondji CS, David JP, Hawkes NJ, Lumjuan N, Nelson DR, Drane DR, Karunaratne SH, Hemingway J, Black WC, Ranson H. Genomic analysis of detoxification genes in the mosquito Aedes aegypti . Insect Biochem Mol Biol 2008; 38: 113-123
- 117 Hurst R, Bao Y, Jemth P, Mannervik B, Williamson G. Phospholipid hydroperoxide glutathione peroxidase activity of human glutathione transferases. Biochem J 1998; 332 (Pt. 1): 97-100
- 118 Lumjuan N, McCarroll L, Prapanthadara LA, Hemingway J, Ranson H. Elevated activity of an Epsilon class glutathione transferase confers DDT resistance in the dengue vector, Aedes aegypti . Insect Biochem Mol Biol 2005; 35: 861-871
- 119 Ranson H, Claudianos C, Ortelli F, Abgrall C, Hemingway J, Sharakhova MV, Unger MF, Collins FH, Feyereisen R. Evolution of supergene families associated with insecticide resistance. Science 2002; 298: 179-181
- 120 Enayati AA, Ranson H, Hemingway J. Insect glutathione transferases and insecticide resistance. Insect Mol Biol 2005; 14: 3-8
- 121 Lumjuan N, Stevenson BJ, Prapanthadara LA, Somboon P, Brophy PM, Loftus BJ, Severson DW, Ranson H. The Aedes aegypti glutathione transferase family. Insect Biochem Mol Biol 2007; 37: 1026-1035
- 122 Lee HL, Chong WL. Glutathion S-transferase activity and DDT-susceptibility of Malaysian mosquitos. Southeast Asian J Trop Med Public Health 1995; 26: 164-167
- 123 Prapanthadara LA, Promtet N, Koottathep S, Somboon P, Suwonkerd W, McCarroll L, Hemingway J. Mechanisms of DDT and permethrin resistance in Aedes aegypti from Chiang Mai, Thailand. Dengue Bull 2002; 26: 185-189
- 124 Ranson H, Rossiter L, Ortelli F, Jensen B, Wang X, Roth CW, Collins FH, Hemingway J. Identification of a novel class of insect glutathione S-transferases involved in resistance to DDT in the malaria vector Anopheles gambiae . Biochem J 2001; 359: 295-304
- 125 Wongtrakul J, Pongjaroenkit S, Leelapat P, Nachaiwieng W, Prapanthadara LA, Ketterman AJ. Expression and characterization of three new glutathione transferases, an epsilon (AcGSTE2-2), omega (AcGSTO1-1), and theta (AcGSTT1-1) from Anopheles cracens (Diptera: Culicidae), a major Thai malaria vector. J Med Entomol 2010; 47: 162-171
- 126 Kostaropoulos I, Papadopoulos AI, Metaxakis A, Boukouvala E, Papadopoulou-Mourkidou E. The role of glutathione S-transferases in the detoxification of some organophosphorus insecticides in larvae and pupae of the yellow mealworm, Tenebrio molitor (Coleoptera: Tenebrionidae). Pest Manag Sci 2001; 57: 501-508
- 127 Vontas J, Kioulos E, Pavlidi N, Morou E, della Torre A, Ranson H. Insecticide resistance in the major dengue vectors Aedes albopictus and Aedes aegypti . Pestic Biochem Phys 2012; 104: 126-131
- 128 Armstrong R. Structure, catalytic mechanism, and evolution of the glutathione S-transferase. Chem Res Toxicol 1997; 10: 2-18
- 129 Ketterman AJ, Saisawang C, Wongsantichon J. Insect glutathione transferases. Drug Metab Rev 2011; 43: 253-265
- 130 Ritter K, Nes W. The effects of cholesterol on the development of Helio thiszea . J Insect Physiol 1981; 27: 175-182
- 131 Grieneisen ML, Warren JT, Gilbert LI. Early steps in ecdysteroid biosynthesis: evidence for the involvement of cytochrome P-450 enzymes. Insect Biochem Mol Biol 1993; 23: 13-23
- 132 Zdobnov EM, von Mering C, Letunic I, Torrents D, Suyama M, Copley RR, Christophides GK, Thomasova D, Holt RA, Subramanian GM, Mueller HM, Dimopoulos G, Law JH, Wells MA, Birney E, Charlab R, Halpern AL, Kokoza E, Kraft CL, Lai Z, Lewis S, Louis C, Barillas-Mury C, Nusskern D, Rubin GM, Salzberg SL, Sutton GG, Topalis P, Wides R, Wincker P, Yandell M, Collins FH, Ribeiro J, Gelbart WM, Kafatos FC, Bork P. Comparative genome and proteome analysis of Anopheles gambiae and Drosophila melanogaster . Science 2002; 298: 149-159
- 133 Kim MS, Wessely V, Lan Q. Identification of mosquito sterol carrier protein-2 inhibitors. J Lipid Res 2005; 46: 650-657
- 134 Peng R, Maklokova VI, Chandrashekhar JH, Lan Q. In vivo functional genomic studies of sterol carrier protein-2 gene in the yellow fever mosquito. PLoS One 2011; 6: e18030
- 135 Gong J, Hou Y, Zha XF, Lu C, Zhu Y, Xia QY. Molecular cloning and characterization of Bombyx mori sterol carrier protein x/sterol carrier protein 2 (SCPx/SCP2) gene. DNA Seq 2006; 17: 326-333
- 136 Guo XR, Zheng SC, Liu L, Feng QL. The sterol carrier protein 2/3-oxoacyl-CoA thiolase (SCPx) is involved in cholesterol uptake in the midgut of Spodoptera litura: gene cloning, expression, localization and functional analyses. BMC Mol Biol 2009; 10: 102
- 137 Renshaw PF, Janoff AS, Miller KW. On the nature of dilute aqueous cholesterol suspensions. J Lipid Res 1983; 24: 47-51
- 138 Arrese EL, Canavoso LE, Jouni ZE, Pennington JE, Tsuchida K, Wells MA. Lipid storage and mobilization in insects: current status and future directions. Insect Biochem Mol Biol 2001; 31: 7-17
- 139 Schroeder F, Gallegos AM, Atshaves BP, Storey SM, McIntosh AL, Petrescu AD, Huang H, Starodub O, Chao H, Yang H, Frolov A, Kier AB. Recent advances in membrane microdomains: rafts, caveolae, and intracellular cholesterol trafficking. Exp Biol Med (Maywood) 2001; 226: 873-890
- 140 Seedorf U, Ellinghaus P, Roch Nofer J. Sterol carrier protein-2. Biochim Biophys Acta 2000; 1486: 45-54
- 141 Vyazunova I, Wessley V, Kim M, Lan Q. Identification of two sterol carrier protein-2 like genes in the yellow fever mosquito, Aedes aegypti . Insect Mol Biol 2007; 16: 305-314
- 142 Krebs KC, Lan Q. Isolation and expression of a sterol carrier protein-2 gene from the yellow fever mosquito, Aedes aegypti . Insect Mol Biol 2003; 12: 51-60
- 143 Lan Q, Massey RJ. Subcellular localization of the mosquito sterol carrier protein-2 and sterol carrier protein-x. J Lipid Res 2004; 45: 1468-1474
- 144 Lan Q, Wessely V. Expression of a sterol carrier protein-x gene in the yellow fever mosquito, Aedes aegypti . Insect Mol Biol 2004; 13: 519-529
- 145 Dyer DH, Wessely V, Forest KT, Lan Q. Three-dimensional structure/function analysis of SCP-2-like2 reveals differences among SCP-2 family members. J Lipid Res 2008; 49: 644-653
Correspondence
Publikationsverlauf
Eingereicht: 26. Juni 2020
Angenommen nach Revision: 21. November 2020
Artikel online veröffentlicht:
21. Dezember 2020
© 2020. Thieme. All rights reserved.
Georg Thieme Verlag KG
Rüdigerstraße 14, 70469 Stuttgart, Germany
-
References
- 1 Newman DJ, Cragg GM. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J Nat Prod 2020; 83: 770-803
- 2 Saldivar-Gonzalez FI, Valli M, Andricopulo AD, da Silva Bolzani V, Medina-Franco JL. Chemical space and diversity of the NuBBE database: A chemoinformatic characterization. J Chem Inf Model 2019; 59: 74-85
- 3 Myers N, Mittermeier RA, Mittermeier CG, da Fonseca GA, Kent J. Biodiversity hotspots for conservation priorities. Nature 2000; 403: 853-858
- 4 Eiten G. Cerrado: Caracterização, Ocupação e Perspectivas. In: Pinto MN. ed. Vegetação do Cerrado. Brasília: Editora da Universidade de Brasília; 1994: 17-73
- 5 Simon MF, Grether R, de Queiroz LP, Skema C, Pennington RT, Hughes CE. Recent assembly of the Cerrado, a neotropical plant diversity hotspot, by in situ evolution of adaptations to fire. Proc Natl Acad Sci U S A 2009; 106: 20359-20364
- 6 Machado R, Neto M, Pereira P, Caldas E, Gonçalves D, Santos N, Tabor K, Steininger M. Estimativas de perda da área do Cerrado brasileiro. Brasília, DF: Conservação Internacional; 2004
- 7 Flausino O, Valli M, Bolzani V. Biodiversidade brasileira: uma fonte potencial de agentes terapêuticos ainda inexplorada. In: Yunes R, Cechinel Filho V. eds. Química de produtos naturais: novos fármacos e a moderna farmacognosia. 3rd ed.. Itajaí: Univali; 2012: 353-383
- 8 Pilon AC, Valli M, Dametto AC, Pinto MEF, Freire RT, Castro-Gamboa I, Andricopulo AD, Bolzani VS. NuBBEDB: an updated database to uncover chemical and biological information from Brazilian biodiversity. Sci Rep 2017; 7: 7215
- 9 Pierson TC, Diamond MS. The continued threat of emerging flaviviruses. Nat Microbiol 2020; 5: 796-812
- 10 Sinka ME, Bangs MJ, Manguin S, Rubio-Palis Y, Chareonviriyaphap T, Coetzee M, Mbogo CM, Hemingway J, Patil AP, Temperley WH, Gething PW, Kabaria CW, Burkot TR, Harbach RE, Hay SI. A global map of dominant malaria vectors. Parasit Vectors 2012; 5: 69
- 11 Dunphy BM, Kovach KB, Gehrke EJ, Field EN, Rowley WA, Bartholomay LC, Smith RC. Long-term surveillance defines spatial and temporal patterns implicating Culex tarsalis as the primary vector of West Nile virus. Sci Rep 2019; 9: 6637
- 12 Leparc-Goffart I, Nougairede A, Cassadou S, Prat C, de Lamballerie X. Chikungunya in the Americas. Lancet 2014; 383: 514
- 13 Kraemer MU, Sinka ME, Duda KA, Mylne AQ, Shearer FM, Barker CM, Moore CG, Carvalho RG, Coelho GE, Van Bortel W, Hendrickx G, Schaffner F, Elyazar IR, Teng HJ, Brady OJ, Messina JP, Pigott DM, Scott TW, Smith DL, Wint GR, Golding N, Hay SI. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus . Elife 2015; 4: e08347
- 14 Wilder-Smith A, Ooi EE, Horstick O, Wills B. Dengue. Lancet 2019; 393: 350-363
- 15 WHO. Dengue and severe dengue. Accessed May 30, 2020 at: https://www.who.int/news-room/fact-sheets/detail/dengue-and-severe-dengue
- 16 Brady OJ, Gething PW, Bhatt S, Messina JP, Brownstein JS, Hoen AG, Moyes CL, Farlow AW, Scott TW, Hay SI. Refining the global spatial limits of dengue virus transmission by evidence-based consensus. PLoS Negl Trop Dis 2012; 6: e1760
- 17 Bhatt S, Gething PW, Brady OJ, Messina JP, Farlow AW, Moyes CL, Drake JM, Brownstein JS, Hoen AG, Sankoh O, Myers MF, George DB, Jaenisch T, Wint GR, Simmons CP, Scott TW, Farrar JJ, Hay SI. The global distribution and burden of dengue. Nature 2013; 496: 504-507
- 18 PAHO. Casos reportados de dengue en las Américas. Accessed May 30, 2020 at: https://www.paho.org/data/index.php/es/temas/indicadores-dengue/dengue-nacional/9-dengue-pais-ano.html
- 19 WHO. Zika virus. Accessed May 30, 2020 at: https://www.who.int/en/news-room/fact-sheets/detail/zika-virus
- 20 Brasil P, Pereira JP, Moreira ME, Ribeiro Nogueira RM, Damasceno L, Wakimoto M, Rabello RS, Valderramos SG, Halai UA, Salles TS, Zin AA, Horovitz D, Daltro P, Boechat M, Raja Gabaglia C, Carvalho de Sequeira P, Pilotto JH, Medialdea-Carrera R, Cotrim da Cunha D, Abreu de Carvalho LM, Pone M, Machado Siqueira A, Calvet GA, Rodrigues Baiao AE, Neves ES, Nassar de Carvalho PR, Hasue RH, Marschik PB, Einspieler C, Janzen C, Cherry JD, Bispo de Filippis AM, Nielsen-Saines K. Zika virus infection in pregnant women in Rio de Janeiro. N Engl J Med 2016; 375: 2321-2334
- 21 WHO. Chikungunya. Accessed May 30, 2020 at: https://www.who.int/news-room/fact-sheets/detail/chikungunya
- 22 Guy B, Briand O, Lang J, Saville M, Jackson N. Development of the Sanofi Pasteur tetravalent dengue vaccine: One more step forward. Vaccine 2015; 33: 7100-7111
- 23 Guy B, Jackson N. Dengue vaccine: hypotheses to understand CYD-TDV-induced protection. Nat Rev Microbiol 2016; 14: 45-54
- 24 WHO. Questions and Answers on Dengue Vaccines. Accessed May 30, 2020 at: https://www.who.int/immunization/research/development/dengue_q_and_a/en/
- 25 WHO. Neglected tropical diseases. Accessed May 30, 2020 at: https://www.who.int/neglected_diseases/en/
- 26 Barbosa PBB, Oliveira JM, Chagas JM, Rabelo LMA, Medeiros GF, Giodani RB, Silva EA, Uchôa AF, Ximenes MFF. Evaluation of seed extracts from plants found in the Caatinga biome for the control of Aedes aegypti . Parasitol Res 2014; 113: 3565-3580
- 27 Ministério da Saúde. Fundação Nacional de Saúde. Manual de vigilância epidemiológica de febre amarela. Brasília, DF: Ministério da Saúde: Fundação Nacional de Saúde; 2004. Accessed December 15 2020 at: https://bvsms.saude.gov.br/bvs/publicacoes/manual_vigilancia_epid_febre_amarela.pdf
- 28 Porto KR, Roel AR, Silva MM, Coelho RM, Scheleder EJ, Jeller AH. Atividade larvicida do óleo de Anacardium humile Saint Hill sobre Aedes aegypti (Linnaeus, 1762) (Diptera, Culicidae). Rev Bras Med Trop 2008; 41: 586-589
- 29 de Mendonca FA, da Silva KF, dos Santos KK, Ribeiro Júnior KA, SantʼAna AE. Activities of some Brazilian plants against larvae of the mosquito Aedes aegypti . Fitoterapia 2005; 76: 629-636
- 30 de Carvalho GHF, de Andrade MA, de Araujo CN, Santos ML, de Castro NA, Charneau S, Monnerat R, de Santana JM, Bastos IMD. Larvicidal and pupicidal activities of eco-friendly phenolic lipid products from Anacardium occidentale nutshell against arbovirus vectors. Environ Sci Pollut Res Int 2019; 26: 5514-5523
- 31 Souza TM, Farias DF, Soares BM, Viana MP, Lima GPG, Machado LKA, Morais SM, Carvalho AFU. Toxicity of Brazilian plant seed extracts to two strains of Aedes aegypti (Diptera: Culicidae) and nontarget animals. J Med Entomol 2011; 48: 846-851
- 32 Napoleao TH, Pontual EV, de Albuquerque Lima T, de Lima Santos ND, Sá RA, Coelho LC, do Amaral Ferraz Navarro DM, Paiva PM. Effect of Myracrodruon urundeuva leaf lectin on survival and digestive enzymes of Aedes aegypti larvae. Parasitol Res 2012; 110: 609-616
- 33 Silva AG, Almeida DL, Ronchi SN, Bento AC, Scherer R, Ramos AC, Cruz ZM. The essential oil of Brazilian pepper, Schinus terebinthifolia Raddi in larval control of Stegomyia aegypti (Linnaeus, 1762). Parasit Vectors 2010; 3: 79
- 34 Santos CCS, Araujo SS, Santos ALLM, Almeida ECV, Dias AS, Damascena NP, Santos DM, Santos MIS, Júnior KALR, Pereira CKB, Lima ACB, Shan AYKV, Santʼana AEG, Estevam CS, Araujo BS. Evaluation of the toxicity and molluscicidal and larvicidal activities of Schinopsis brasiliensis stem bark extract and its fractions. Rev Bras Farmacogn 2014; 24: 298-303
- 35 de Omena MC, Navarro DMAF, de Paula JE, Luna JS, de Lima MRF, SantʼAna AEG. Larvicidal activities against Aedes aegypti of some Brazilian medicinal plants. Bioresour Technol 2007; 98: 2549-2556
- 36 Grzybowski A, Tiboni M, Silva MA, Chitolina RF, Passos M, Fontana JD. Synergistic larvicidal effect and morphological alterations induced by ethanolic extracts of Annona muricata and Piper nigrum against the dengue fever vector Aedes aegypti . Pest Manag Sci 2013; 69: 589-601
- 37 Rodrigues AM, Silva AAS, Pinto CCC, Lima Dos Santos D, Carneiro de Freitas JC, Martins VEP, Maia de Morais S. Larvicidal and enzymatic inhibition effects of Annona muricata seed extract and main constituent annonacin against Aedes aegypti and Aedes albopictus (Diptera: Culicidae). Pharmaceuticals (Basel) 2019; 12: 112
- 38 Simas NK, Dellamora EDL, Schripsema J, Lage CLS, de Oliveira AM, Wessjohann L, Porzel A, Kuster RM. Acetylenic 2-phenylethylamides and new isobutylamides from Acmella oleracea (L.) R. K. Jansen, a Brazilian spice with larvicidal activity on Aedes aegypti . Phytochem Lett 2013; 6: 67-72
- 39 Lavor PL, Santiago GMP, Gois RWD, de Sousa LM, Bezerra GD, Romero NR, Arriaga AMC, Lemos TLG, Alves PB, Gomes PCS. Larvicidal activity against Aedes aegypti of essential oils from northeast Brazil. Nat Prod Commun 2012; 7: 1391-1392
- 40 Botas GDS, Cruz RAS, de Almeida FB, Duarte JL, Araujo RS, Souto RNP, Ferreira R, Carvalho JCT, Santos MG, Rocha L, Pereira VLP, Fernandes CP. Baccharis reticularia DC. and limonene nanoemulsions: Promising larvicidal agents for Aedes aegypti (Diptera: Culicidae) control. Molecules 2017; 22: 1990
- 41 Marques MM, Morais SM, Vieira IG, Vieira MG, Raquel A, Silva A, De Almeida RR, Guedes MI. Larvicidal activity of Tagetes erecta against Aedes aegypti . J Am Mosq Control Assoc 2011; 27: 156-158
- 42 Garcez WS, Garcez FR, da Silva LMGE, Hamerski L. Larvicidal activity against Aedes aegypti of some plants native to the West-Central region of Brazil. Bioresour Technol 2009; 100: 6647-6650
- 43 Borges JCM, Silva EAP e, de Barros TCA, Soares IM, Ascêncio SD, Fidelis RR, de Souza Aguiar RW. Chemical composition, oviposition deterrent and larvicidal activities of the wood extracts of Tabebuia avellanedae from the Cerrado of Brazil. J Med Plant Res 2018; 12: 404-414
- 44 Costa JGM, Pessoa ODL, Menezes EA, Santiago GMP, Lemos TLG. Composition and larvicidal activity of essential oils from heartwood of Auxemma glazioviana Taub. (Boraginaceae). Flavour Frag J 2004; 19: 529-531
- 45 Santos RP, Nunes EP, Nascimento RF, Santiago GMP, Menezes GHA, Silveira ER, Pessoa ODL. Chemical composition and larvicidal activity of the essential oils of Cordia leucomalloides and Cordia curassavica from the northeast of Brazil. J Brazil Chem Soc 2006; 17: 1027-1030
- 46 de Menezes JESA, Lemos TLG, Silveira ER, Pessoa ODL, Santiago GMP. Chemical composition and larvicidal activity of the essential oil from leaves of Cordia globosa (Jacq.) HBK from northeastern Brazil. J Essent Oil Res 2006; 18: 253-255
- 47 da Silva RC, Milet-Pinheiro P, Bezerra da Silva PC, da Silva AG, da Silva MV, Navarro DM, da Silva NH. (E)-caryophyllene and alpha-humulene: Aedes aegypti oviposition deterrents elucidated by gas chromatography-electrophysiological assay of Commiphora leptophloeos leaf oil. PLoS One 2015; 10: e0144586
- 48 Oliveira PV, Ferreira JC, Moura FS, Lima GS, de Oliveira FM, Oliveira PE, Conserva LM, Giulietti AM, Lemos RP. Larvicidal activity of 94 extracts from ten plant species of northeastern of Brazil against Aedes aegypti L. (Diptera: Culicidae). Parasitol Res 2010; 107: 403-407
- 49 de Lima GP, de Souza TM, de Paula Freire G, Farias DF, Cunha AP, Ricardo NM, de Morais SM, Carvalho AF. Further insecticidal activities of essential oils from Lippia sidoides and Croton species against Aedes aegypti L. Parasitol Res 2013; 112: 1953-1958
- 50 Morais SM, Cavalcanti ESB, Bertini LM, Oliveira CLL, Rodrigues JRB, Cardoso JHL. Larvicidal activity of essential oils from Brazilian Croton species against Aedes aegypti L. J Am Mosq Control Assoc 2006; 22: 161-164
- 51 Doria GA, Silva WJ, Carvalho GA, Alves PB, Cavalcanti SC. A study of the larvicidal activity of two Croton species from northeastern Brazil against Aedes aegypti . Pharm Biol 2010; 48: 615-620
- 52 Santos GK, Dutra KA, Lira CS, Lima BN, Napoleao TH, Paiva PM, Maranhao CA, Brandao SS, Navarro DM. Effects of Croton rhamnifolioides essential oil on Aedes aegypti oviposition, larval toxicity and trypsin activity. Molecules 2014; 19: 16573-16587
- 53 Torres MC, Assuncao JC, Santiago GM, Andrade-Neto M, Silveira ER, Costa-Lotufo LV, Bezerra DP, Marinho Filho JD, Viana FA, Pessoa OD. Larvicidal and nematicidal activities of the leaf essential oil of Croton regelianus . Chem Biodivers 2008; 5: 2724-2728
- 54 Carvalho Kda S, Silva SL e, de Souza IA, Gualberto SA, da Cruz RC, Dos Santos FR, de Carvalho MG. Toxicological evaluation of essential oil from the leaves of Croton tetradenius (Euphorbiaceae) on Aedes aegypti and Mus musculus . Parasitol Res 2016; 115: 3441-3448
- 55 Farias DF, Cavalheiro MG, Viana MP, Queiroz VA, Rocha-Bezerra LCB, Vasconcelos IM, Morais SM, Carvalho AFU. Water extracts of Brazilian leguminous seeds as rich sources of larvicidal compounds against Aedes aegypti L. An Acad Bras Ciênc 2010; 82: 585-594
- 56 Trindade FTT, Stabeli RG, Pereira AA, Facundo VA, Silva ADE. Copaifera multijuga ethanolic extracts, oil-resin, and its derivatives display larvicidal activity against Anopheles darlingi and Aedes aegypti (Diptera: Culicidae). Rev Bras Farmacogn 2013; 23: 464-470
- 57 Dalarmi L, da Silva CB, Ocampos FMM, Burci LM, do Nascimento KF, de Jesus C, Dias JdFG, Miguel MD, Miguel OG, Zanin SMW. Larvicidal activity of Dalbergia brasiliensis (Fabaceae – Papilionoideae) on Aedes aegypti . Afr J Pharm Pharmaco 2018; 9: 881-885
- 58 Gusmao DS, Pascoa V, Mathias L, Curcino Vieira IJ, Braz-Filho R, Alves Lemos FJ. Derris (Lonchocarpus) urucu (Leguminosae) extract modifies the peritrophic matrix structure of Aedes aegypti (Diptera: Culicidae). Mem Inst Oswaldo Cruz 2002; 97: 371-375
- 59 Aguiar JC, Santiago GM, Lavor PL, Veras HN, Ferreira YS, Lima MA, Arriaga AM, Lemos TL, Lima JQ, de Jesus HC, Alves PB, Braz-Filho R. Chemical constituents and larvicidal activity of Hymenaea courbaril fruit peel. Nat Prod Commun 2010; 5: 1977-1980
- 60 Porto KRA, Motti PR, Yano M, Roel AR, Cardoso CAL, Matias R. Screening of plant extracts and fractions on Aedes aegypti larvae found in the state of Mato Grosso do Sul (linnaeus, 1762) (culicidae). An Acad Bras Ciênc 2017; 8: 895-906
- 61 Arriaga AM, Lima JQ, Vasconcelos JN e, de Oliveira MC, Lemos TL, Fonseca AM, Malcher GT, Santiago GM, Mafezoli J, Braz-Filho R. Antioxidant and larvicidal activities of Tephrosia egregia Sandw against Aedes aegypti . Nat Prod Commun 2009; 4: 529-530
- 62 Costa JGM, Rodrigues FFG, Angélico EC, Silva MR, Mota ML, Santos NKA, Cardoso ALH, Lemos TLG. Estudo químico-biológico dos óleos essenciais de Hyptis martiusii, Lippia sidoides e Syzigium aromaticum frente às larvas do Aedes aegypti . Rev Bras Farmacogn 2005; 15: 304-309
- 63 Silva WJ, Doria GA, Maia RT, Nunes RS, Carvalho GA, Blank AF, Alves PB, Marcal RM, Cavalcanti SC. Effects of essential oils on Aedes aegypti larvae: alternatives to environmentally safe insecticides. Bioresour Technol 2008; 99: 3251-3255
- 64 Ricarte LP, Bezerra GP, Romero NR, Silva HCD, Lemos TLG, Arriaga AMC, Alves PB, Santos MBD, Militao GCG, Silva TDS, Braz-Filho R, Santiago GMP. Chemical composition and biological activities of the essential oils from Vitex-agnus castus, Ocimum campechianum and Ocimum carnosum . An Acad Bras Cienc 2020; 92: e20180569
- 65 Azevedo SG, Mar JM, da Silva LS, Franca LP, Machado MB, Tadei WP, Bezerra JD, dos Santos AL, Sanches EA. Bioactivity of Licaria puchury-major essential oil against Aedes aegypti, Tetranychus urticae and Cerataphis lataniae . Rec Nat Prod 2018; 12: 229-239
- 66 da Silva HHG, Geris R, Rodrigues E, Rocha C, da Silva IG. Larvicidal activity of oil-resin fractions from the Brazilian medicinal plant Copaifera reticulata Ducke (Leguminosae-Caesalpinoideae) against Aedes aegypti (Diptera, Culicidae). Rev Soc Bras Med Trop 2007; 40: 264-267
- 67 Gois RWD, de Sousa LM, Lemos TLG, Arriaga AMC, Andrade-Neto M, Santiago GMP, Ferreira YS, Alves PB, de Jesus HCR. Chemical composition and larvicidal effects of essential oil from Bauhinia acuruana (Moric) againstAedes aegypti. . J Essent Oil Res 2011; 23: 59-62
- 68 dos Santos EA, de Carvalho CM, Costa ALS, Conceicao AS, Moura FDP, Santana AEG. Bioactivity evaluation of plant extracts used in indigenous medicine against the snail, Biomphalaria glabrata, and the larvae of Aedes aegypti . Evid Based Complement Alternat 2012; 2012: 846583
- 69 Ferreira MDL, Fernandes DA, Nunes FC, Teles YCF, Rolim YM, da Silva CM, de Albuquerque JBL, Agra MF, de Souza MFV. Phytochemical study of Waltheria viscosissima and evaluation of its larvicidal activity against Aedes aegypti . Rev Bras Farmacogn 2019; 29: 582-590
- 70 Ferreira MR, Santiago RR, de Souza TP, Egito ES, Oliveira EE, Soares LA. Development and evaluation of emulsions from Carapa guianensis (Andiroba) oil. AAPS PharmSciTech 2010; 11: 1383-1390
- 71 Sarmento UC, Miguita CH, Almeida LH, Gaban CR, Silva LM, Souza AS, Garcez WS, Garcez FR. Larvicidal efficacies of plants from Midwestern Brazil: melianodiol from Guarea kunthiana as a potential biopesticide against Aedes aegypti . Mem Inst Oswaldo Cruz 2016; 111: 469-474
- 72 Magalhaes LAM, Lima MD, Marques MOM, Facanali R, Pinto ACD, Tadei WP. Chemical composition and larvicidal activity against Aedes aegypti larvae of essential oils from four Guarea species. Molecules 2010; 15: 5734-5741
- 73 da Silva AG, Alves RCC, Filho CMB, Bezerra-Silva PC, dos Santos LMM, Foglio MA, Navarro DMDF, da Silva MV, Correia MTD. Chemical composition and larvicidal activity of the essential oil from leaves of Eugenia brejoensis Mazine (Myrtaceae). J Essent Oil Bear Pl 2015; 18: 1441-1447
- 74 Neves ID, Rezende SRD, Kirk JM, Pontes EG, De Carvalho MG. Composition and larvicidal activity of essential oil of Eugenia candolleana DC. (MYRTACEAE) against Aedes aegypti . Rev Virtual Quim 2017; 9: 2305-2315
- 75 Dias CN, Alves LP, Rodrigues KA, Brito MC, Rosa Cdos S, do Amaral FM, Monteiro Odos S, Andrade EH, Maia JG, Moraes DF. Chemical composition and larvicidal activity of essential oils extracted from Brazilian legal amazon plants against Aedes aegypti L. (Diptera: Culicidae). Evid Based Complement Alternat Med 2015; 2015: 490765
- 76 Rosa CS, Veras KS, Silva PR, Lopes Neto JJ, Cardoso HLM, Alves LPL, Brito MCA, Amaral FMM, Maia JGS, Monteiro OS, Moraes DFC. Composição química e toxicidade frente Aedes aegypti L. e Artemia salina Leach do óleo essencial das folhas de Myrcia sylvatica (G. Mey.) DC. Rev Bras Pl Med 2016; 18: 19-26
- 77 Mendes LA, Martins GF, Valbon WR, de Souza TD, Menini L, Ferreira A, Ferreira MFD. Larvicidal effect of essential oils from Brazilian cultivars of guava on Aedes aegypti L. Ind Crop Prod 2017; 108: 684-689
- 78 Kanis LA, Antonio RD, Antunes EP, Prophiro JS, da Silva OS. Larvicidal effect of dried leaf extracts from Pinus caribaea against Aedes aegypti (Linnaeus, 1762) (Diptera: Culicidae). Rev Soc Bras Med Trop 2009; 42: 373-376
- 79 Oliveira GL, Cardoso SK, Lara CR, Vieira TM, Guimaraes EF, Figueiredo LS, Martins ER, Moreira DL, Kaplan MA. Chemical study and larvicidal activity against Aedes aegypti of essential oil of Piper aduncum L. (Piperaceae). An Acad Bras Cienc 2013; 85: 1227-1234
- 80 da Silva MF, Bezerra-Silva PC, de Lira CS, de Lima Albuquerque BN, Agra Neto AC, Pontual EV, Maciel JR, Paiva PM, Navarro DM. Composition and biological activities of the essential oil of Piper corcovadensis (Miq.) C. DC (Piperaceae). Exp Parasitol 2016; 165: 64-70
- 81 do Nascimento JC, David JM, Barbosa LC, de Paula VF, Demuner AJ, David JP, Conserva LM, Ferreira JC, Guimaraes EF. Larvicidal activities and chemical composition of essential oils from Piper klotzschianum (Kunth) C. DC. (Piperaceae). Pest Manag Sci 2013; 69: 1267-1271
- 82 Autran ES, Neves IA, da Silva CSB, Santos GKN, da Camara CAG, Navarro DMAF. Chemical composition, oviposition deterrent and larvicidal activities against Aedes aegypti of essential oils from Piper marginatum Jacq. (Piperaceae). Bioresour Technol 2009; 100: 2284-2288
- 83 Marques AM, Velozo LS, Carvalho MA, Serdeiro MT, Honorio NA, Kaplan MAC, Maleck M. Larvicidal activity of Ottonia anisum metabolites against Aedes aegypti: A potential natural alternative source for mosquito vector control in Brazil. J Vector Dis 2017; 54: 61-68
- 84 Cavalcanti ESB, de Morais SM, Lima MAA, Santana EWP. Larvicidal activity of essential oils from Brazilian plants against Aedes aegypti L. Mem Inst Oswaldo Cruz 2004; 99: 541-544
- 85 Arriaga AMC, Rodrigues FEA, Lemos TLG, de Oliveira MDF, Lima JQ, Santiago GMP, Braz-Filho R, Matezoli J. Composition and larvicidal activity of essential oil from Stemodia maritima L. Nat Prod Commun 2007; 2: 1237-1239
- 86 Ferreira RMDA, Dʼhaveloose NP, Cruz RAS, Araújo RS, Carvalho JCT, Rocha L, Fernandes LP, Da Costa TS, Fernandes CP, Souto RNP. Nano-emulsification enhances the larvicidal potential of the essential oil of Siparuna guianensis (Laurales: Siparunaceae) against Aedes (Stegomyia) aegypti (Diptera: Culicidae). J Med Entomol 2020; 57: 788-796
- 87 Avelar Amado P, Fonseca Castro AH, Nunes Alves S, Brentan Silva D, Alexandre Carollo C. Alves Rodrigues Dos Santos Lima L. Phenolic compounds: antioxidant and larvicidal potential of Smilax brasiliensis Sprengel leaves. Nat Prod Res 2019; 17: 2545-2553
- 88 Santiago GMP, Lemos TLG, Pessoa ODL, Arriaga AMC, Matos FJA, Lima MAS, Santos HS, Lima DCL, Barbosa FG, Luciano JHS, Silveira ER, de Menezes GHA. Larvicidal activity against Aedes aegypti L. (Diptera: Culicidae) of essential oils of Lippia species from Brazil. Nat Prod Commun 2006; 1: 573-576
- 89 Nascimento AMD, Maia TDS, Soares TES, Menezes LRA, Scher R, Costa EV, Cavalcanti SCH, La Corte R. Repellency and larvicidal activity of essential oils from Xylopia laevigata, Xylopia frutescens, Lippia pedunculosa, and their individual compounds against Aedes aegypti Linnaeus. Neotrop Entomol 2017; 46: 223-230
- 90 Costa M, Pereira M, Oliveira S, Souza P, Dallʼoglio E, Alves T. Anonáceas provocam mortalidade em larvas de Aedes aegypti (Linnaeus, 1762) (Dipteria: Culidae). Rev Bras Biocienc 2013; 11: 184-190
- 91 Costa MS, Santana AE, Oliveira LL, Zanuncio JC, Serrao JE. Toxicity of squamocin on Aedes aegypti larvae, its predators and human cells. Pest Manag Sci 2017; 73: 636-640
- 92 De Sousa FDM, Grossi SM, Monteiro GC, Demarque DP, Espindola LS. Dereplication and isolation of larvicidal compounds from annonaceae species against Aedes aegypti . Rev Bras Farmacogn 2020; 30: 123-126
- 93 Rodrigues AMS, De Paula JE, Degallier N, Molez JE, Espindola LS. Larvicidal activity of some Cerrado plant extracts against Aedes aegypti . J Am Mosq Control Assoc 2006; 22: 314-317
- 94 Borges RA, Cavasin GM, Silva IG, Arruda W, Oliveira ESF, Silva HHG, Martins F. Mortalidade e alterações morfológicas provocadas pela ação inibidora do diflubenzuron na ecdise de larvas de Aedes aegypti (Diptera, Culicidae). Rev Patol Trop 2004; 33: 91-105
- 95 Barreto CF, Cavasin GM, Silva HHG, Silva IG. Estudo das alterações morfo-histológicas em larvas de Aedes aegypti (Diptera, Culicidae) submetidas ao extrato bruto etanólico de Sapindus saponaria Lin (Sapindaceae). Rev Patol Trop 2006; 35: 37-57
- 96 Correia SJ, David JP, David JM. Secundary metabolites from species of Anacardiaceae. Quim Nova 2006; 29: 1287-1300
- 97 Audi EA, Otobone F, Martins JV, Cortez DA. Preliminary evaluation of Kielmeyera coriacea leaves extract on the central nervous system. Fitoterapia 2002; 73: 517-519
- 98 Coelho AAM, Paula JE, Espindola LS. Atividade larvicida de extratos vegetais sobre Aedes aegypti (L.) (Diptera: Culicidae), em condições de laboratório. Rev Entom Bras 2009; 4: 1-6
- 99 Ma DL, Chan DSH, Leung CH. Molecular docking for virtual screening of natural product databases. Chem Sci 2011; 2: 1656-1665
- 100 Andricopulo AD, Montanari CA. Structure-activity relationships for the design of small-molecule inhibitors. Mini Rev Med Chem 2005; 5: 585-593
- 101 Ferreira LG, Andricopulo AD. From protein structure to small-molecules: recent advances and applications to fragment-based drug discovery. Curr Top Med Chem 2017; 17: 2260-2270
- 102 Ferreira LG, Oliva G, Andricopulo AD. From medicinal chemistry to human health: Current approaches to drug discovery for cancer and neglected tropical diseases. An Acad Bras Cienc 2018; 90: 645-661
- 103 Chakraborty G, Shin J, Nguyen QT, Harikishore A, Baek K, Yoon HS. Solution structure of FK506-binding protein 12 from Aedes aegypti . Proteins 2012; 80: 2476-2481
- 104 Rajan S, Saw KQ, Nguyen QT, Baek K, Yoon HS. High-resolution crystal structure of FKBP12 from Aedes aegypti . Protein Sci 2012; 21: 1080-1084
- 105 Leite NR, Krogh R, Xu W, Ishida Y, Iulek J, Leal WS, Oliva G. Structure of an odorant-binding protein from the mosquito Aedes aegypti suggests a binding pocket covered by a pH-sensitive “Lid”. PLoS One 2009; 4: e8006
- 106 Leal WS. Odorant reception in insects: roles of receptors, binding proteins, and degrading enzymes. Annu Rev Entomol 2013; 58: 373-391
- 107 Li S, Picimbon JF, Ji S, Kan Y, Chuanling Q, Zhou JJ, Pelosi P. Multiple functions of an odorant-binding protein in the mosquito Aedes aegypti . Biochem Biophys Res Commun 2008; 372: 464-468
- 108 Bohbot J, Pitts RJ, Kwon HW, Rutzler M, Robertson HM, Zwiebel LJ. Molecular characterization of the Aedes aegypti odorant receptor gene family. Insect Mol Biol 2007; 16: 525-537
- 109 Kim IH, Pham V, Jablonka W, Goodman WG, Ribeiro JMC, Andersen JF. A mosquito hemolymph odorant-binding protein family member specifically binds juvenile hormone. J Biol Chem 2017; 292: 15329-15339
- 110 Calvo E, Mans BJ, Ribeiro JM, Andersen JF. Multifunctionality and mechanism of ligand binding in a mosquito antiinflammatory protein. Proc Natl Acad Sci U S A 2009; 106: 3728-3733
- 111 Petchampai N, Murillo-Solano C, Isoe J, Pizarro JC, Scaraffia PY. Distinctive regulatory properties of pyruvate kinase 1 from Aedes aegypti mosquitoes. Insect Biochem Mol Biol 2019; 104: 82-90
- 112 Han Q, Robinson H, Ding H, Christensen BM, Li J. Evolution of insect arylalkylamine N-acetyltransferases: structural evidence from the yellow fever mosquito, Aedes aegypti . Proc Natl Acad Sci U S A 2012; 109: 11669-11674
- 113 Han Q, Gao YG, Robinson H, Ding H, Wilson S, Li J. Crystal structures of Aedes aegypti kynurenine aminotransferase. FEBS J 2005; 272: 2198-2206
- 114 Han Q, Robinson H, Gao YG, Vogelaar N, Wilson SR, Rizzi M, Li J. Crystal structures of Aedes aegypti alanine glyoxylate aminotransferase. J Biol Chem 2006; 281: 37175-37182
- 115 Asad S, Hall-Mendelin S, Asgari S. Downregulation of Aedes aegypti chromodomain helicase DNA binding protein 7/Kismet by Wolbachia and its effect on dengue virus replication. Sci Rep 2016; 6: 36850
- 116 Strode C, Wondji CS, David JP, Hawkes NJ, Lumjuan N, Nelson DR, Drane DR, Karunaratne SH, Hemingway J, Black WC, Ranson H. Genomic analysis of detoxification genes in the mosquito Aedes aegypti . Insect Biochem Mol Biol 2008; 38: 113-123
- 117 Hurst R, Bao Y, Jemth P, Mannervik B, Williamson G. Phospholipid hydroperoxide glutathione peroxidase activity of human glutathione transferases. Biochem J 1998; 332 (Pt. 1): 97-100
- 118 Lumjuan N, McCarroll L, Prapanthadara LA, Hemingway J, Ranson H. Elevated activity of an Epsilon class glutathione transferase confers DDT resistance in the dengue vector, Aedes aegypti . Insect Biochem Mol Biol 2005; 35: 861-871
- 119 Ranson H, Claudianos C, Ortelli F, Abgrall C, Hemingway J, Sharakhova MV, Unger MF, Collins FH, Feyereisen R. Evolution of supergene families associated with insecticide resistance. Science 2002; 298: 179-181
- 120 Enayati AA, Ranson H, Hemingway J. Insect glutathione transferases and insecticide resistance. Insect Mol Biol 2005; 14: 3-8
- 121 Lumjuan N, Stevenson BJ, Prapanthadara LA, Somboon P, Brophy PM, Loftus BJ, Severson DW, Ranson H. The Aedes aegypti glutathione transferase family. Insect Biochem Mol Biol 2007; 37: 1026-1035
- 122 Lee HL, Chong WL. Glutathion S-transferase activity and DDT-susceptibility of Malaysian mosquitos. Southeast Asian J Trop Med Public Health 1995; 26: 164-167
- 123 Prapanthadara LA, Promtet N, Koottathep S, Somboon P, Suwonkerd W, McCarroll L, Hemingway J. Mechanisms of DDT and permethrin resistance in Aedes aegypti from Chiang Mai, Thailand. Dengue Bull 2002; 26: 185-189
- 124 Ranson H, Rossiter L, Ortelli F, Jensen B, Wang X, Roth CW, Collins FH, Hemingway J. Identification of a novel class of insect glutathione S-transferases involved in resistance to DDT in the malaria vector Anopheles gambiae . Biochem J 2001; 359: 295-304
- 125 Wongtrakul J, Pongjaroenkit S, Leelapat P, Nachaiwieng W, Prapanthadara LA, Ketterman AJ. Expression and characterization of three new glutathione transferases, an epsilon (AcGSTE2-2), omega (AcGSTO1-1), and theta (AcGSTT1-1) from Anopheles cracens (Diptera: Culicidae), a major Thai malaria vector. J Med Entomol 2010; 47: 162-171
- 126 Kostaropoulos I, Papadopoulos AI, Metaxakis A, Boukouvala E, Papadopoulou-Mourkidou E. The role of glutathione S-transferases in the detoxification of some organophosphorus insecticides in larvae and pupae of the yellow mealworm, Tenebrio molitor (Coleoptera: Tenebrionidae). Pest Manag Sci 2001; 57: 501-508
- 127 Vontas J, Kioulos E, Pavlidi N, Morou E, della Torre A, Ranson H. Insecticide resistance in the major dengue vectors Aedes albopictus and Aedes aegypti . Pestic Biochem Phys 2012; 104: 126-131
- 128 Armstrong R. Structure, catalytic mechanism, and evolution of the glutathione S-transferase. Chem Res Toxicol 1997; 10: 2-18
- 129 Ketterman AJ, Saisawang C, Wongsantichon J. Insect glutathione transferases. Drug Metab Rev 2011; 43: 253-265
- 130 Ritter K, Nes W. The effects of cholesterol on the development of Helio thiszea . J Insect Physiol 1981; 27: 175-182
- 131 Grieneisen ML, Warren JT, Gilbert LI. Early steps in ecdysteroid biosynthesis: evidence for the involvement of cytochrome P-450 enzymes. Insect Biochem Mol Biol 1993; 23: 13-23
- 132 Zdobnov EM, von Mering C, Letunic I, Torrents D, Suyama M, Copley RR, Christophides GK, Thomasova D, Holt RA, Subramanian GM, Mueller HM, Dimopoulos G, Law JH, Wells MA, Birney E, Charlab R, Halpern AL, Kokoza E, Kraft CL, Lai Z, Lewis S, Louis C, Barillas-Mury C, Nusskern D, Rubin GM, Salzberg SL, Sutton GG, Topalis P, Wides R, Wincker P, Yandell M, Collins FH, Ribeiro J, Gelbart WM, Kafatos FC, Bork P. Comparative genome and proteome analysis of Anopheles gambiae and Drosophila melanogaster . Science 2002; 298: 149-159
- 133 Kim MS, Wessely V, Lan Q. Identification of mosquito sterol carrier protein-2 inhibitors. J Lipid Res 2005; 46: 650-657
- 134 Peng R, Maklokova VI, Chandrashekhar JH, Lan Q. In vivo functional genomic studies of sterol carrier protein-2 gene in the yellow fever mosquito. PLoS One 2011; 6: e18030
- 135 Gong J, Hou Y, Zha XF, Lu C, Zhu Y, Xia QY. Molecular cloning and characterization of Bombyx mori sterol carrier protein x/sterol carrier protein 2 (SCPx/SCP2) gene. DNA Seq 2006; 17: 326-333
- 136 Guo XR, Zheng SC, Liu L, Feng QL. The sterol carrier protein 2/3-oxoacyl-CoA thiolase (SCPx) is involved in cholesterol uptake in the midgut of Spodoptera litura: gene cloning, expression, localization and functional analyses. BMC Mol Biol 2009; 10: 102
- 137 Renshaw PF, Janoff AS, Miller KW. On the nature of dilute aqueous cholesterol suspensions. J Lipid Res 1983; 24: 47-51
- 138 Arrese EL, Canavoso LE, Jouni ZE, Pennington JE, Tsuchida K, Wells MA. Lipid storage and mobilization in insects: current status and future directions. Insect Biochem Mol Biol 2001; 31: 7-17
- 139 Schroeder F, Gallegos AM, Atshaves BP, Storey SM, McIntosh AL, Petrescu AD, Huang H, Starodub O, Chao H, Yang H, Frolov A, Kier AB. Recent advances in membrane microdomains: rafts, caveolae, and intracellular cholesterol trafficking. Exp Biol Med (Maywood) 2001; 226: 873-890
- 140 Seedorf U, Ellinghaus P, Roch Nofer J. Sterol carrier protein-2. Biochim Biophys Acta 2000; 1486: 45-54
- 141 Vyazunova I, Wessley V, Kim M, Lan Q. Identification of two sterol carrier protein-2 like genes in the yellow fever mosquito, Aedes aegypti . Insect Mol Biol 2007; 16: 305-314
- 142 Krebs KC, Lan Q. Isolation and expression of a sterol carrier protein-2 gene from the yellow fever mosquito, Aedes aegypti . Insect Mol Biol 2003; 12: 51-60
- 143 Lan Q, Massey RJ. Subcellular localization of the mosquito sterol carrier protein-2 and sterol carrier protein-x. J Lipid Res 2004; 45: 1468-1474
- 144 Lan Q, Wessely V. Expression of a sterol carrier protein-x gene in the yellow fever mosquito, Aedes aegypti . Insect Mol Biol 2004; 13: 519-529
- 145 Dyer DH, Wessely V, Forest KT, Lan Q. Three-dimensional structure/function analysis of SCP-2-like2 reveals differences among SCP-2 family members. J Lipid Res 2008; 49: 644-653